
’Σ“ΣΘΚΖ÷Ν―ΡΎΚ§κΡΆ®ΙΐΦτΫ”Ζ¥”Π Βœ÷ΕύκΡΤ§ΕΈΒΡΝ§Ϋ”Θ§ ΨΏ”–ΗΏ–ß«“ΈόΚέΒΡΧΊΒψΘ§ ήΒΫΝΥΙψΖΚΙΊΉΔ. ±ΨΈΡΜυ”ΎΖ÷Ν―ΡΎΚ§κΡΒΡΫαΙΙΧΊ’ς”κΦτΫ”Ζ¥”ΠΙΐ≥ΧΘ§ ΫαΚœΫϋΡξά¥ΙΊ”ΎΖ÷Ν―ΡΎΚ§κΡ–‘Ρή”≈Μ·ΚΆ”Π”Ο―–ΨΩΫχ’ΙΫχ––ΝΥΉέΚœΤά ωΘ§ Ϋ“ ΨΤδΉςΈΣ“Μ÷÷»’ΫΞ≥… λΒΡΒΑΑΉ÷ ΙΛ≥ΧΜ·ΦΦ θ‘ΎΒΑΑΉ÷ Μ·―ßΚœ≥…Νλ”ρΒΡ«ΑΨΑΘ§ ≤ΔΦρ“ΣΖ÷ΈωΝΥΡΩ«ΑΖ÷Ν―ΡΎΚ§κΡΙΛΨΏΟφΝΌΒΡΈ Χβ”κΧτ’ΫΘ§ ≤ΔΕ‘Ω…ΡήΒΡΫβΨωΖΫΑΗΫχ––ΝΥ’ΙΆϊ.
Ζ≠“κΚσ–ό ΈΦΪ¥σΒΊΖαΗΜΝΥΒΑΑΉ÷ ΒΡΫαΙΙ”κΙΠΡήΒΡΕύ―υ–‘Θέ1Θί. ’β–©–ό ΈΒΑΑΉ‘Ύ“©ΈοΩΣΖΔΘέ2ΘίΓΔ ΒΑΑΉ÷ Ήι―ßΘέ3Θί“‘ΦΑΒΑΑΉ÷ ΫαΙΙ-ΙΠΡήΙΊœΒΫβΈωΘέ4~8ΘίΒ»ΖΫΟφΨΏ”–÷Ί“ΣΒΡ”Π”Ο. ΡΩ«ΑΘ§ Ζ≠“κΚσ–ό ΈΒΑΑΉΩ…“‘Ά®ΙΐΧλ»ΜΧα»ΓΚΆ…ζΈο±μ¥οΒ»ΖΫΖ®Μώ»Γ. ΒΪ’β–©ΖΫΖ®»‘¥φ‘Ύ“ΜœΒΝ–Έ ΧβΘ§ άΐ»γΘ§ Χλ»ΜΧα»ΓΡ―“‘ΜώΒΟΨυ“Μ–ό ΈΒΡΒΑΑΉ―υΤΖΘ§ Εχ…ζΈο±μ¥οΜώ»ΓΚ§Εύ÷÷Ζ≠“κΚσ–ό ΈΒΡΒΑΑΉ―υΤΖ“ά»Μ¥φ‘ΎάßΡ―Β». ΒΑΑΉ÷ Μ·―ßΚœ≥…ΦΦ θ «ΜώΒΟΖ≠“κΚσ–ό ΈΒΑΑΉΒΡ“Μ÷÷–¬–ΥΦΦ θΘ§ ΗΟΦΦ θΜυ”ΎΙΧœύΕύκΡΚœ≥…ΦΦ θΚΆΕύκΡΤ§ΕΈΝ§Ϋ”ΦΦ θ. ΙΧœύΕύκΡΚœ≥…ΦΦ θΘέ9ΘίΩ…“‘÷±Ϋ”ΫΪΗς÷÷Ζ«Χλ»ΜΑ±ΜυΥαΦΑ¥χ”––ό ΈΜυΆ≈ΒΡΑ±ΜυΥαΖΫ±ψΒΊ“ΐ»κΕύκΡΝ¥÷–ΧΊΕ®ΒΡΈΜΒψ. ΨΓΙήΙΧœύΕύκΡΚœ≥…ΦΦ θΒΡΚœ≥…ΡήΝΠ…œœό“Μ÷±‘Ύ≤ΜΕœΧαΗΏΘέ10ΘίΘ§ ΒΪ¥σ≤ΩΖ÷ΒΑΑΉ»‘»ΜΡ―“‘Ά®ΙΐΙΧœύΚœ≥…“Μ¥Έ–‘Μώ»Γ. “ρ¥ΥΘ§ Ι”ΟΕύκΡΤ§ΕΈΝ§Ϋ”ΦΦ θΡήΙΜΫΪΚ§ΧΊΕ®–ό ΈΒΡΕύκΡΤ§ΕΈΫχ––Ν§Ϋ”Θ§ ΦΪ¥σΒΊά©’ΙΝΥΒΑΑΉ―υΤΖΒΡΜώ»ΓΖΕΈßΘ®ΆΦ1Θ©. Ά®ΙΐΕύκΡΤ§ΕΈΝ§Ϋ”Θ§ ―–ΨΩ’ΏΟ«“―Ψ≠Κœ≥…ΝΥ¥χ”–”ΪΙβ±ξΦ«Θέ11ΘίΘ§ C13ΚΆN15Β»Ά§ΈΜΥΊ±ξΦ«Θέ12ΘίΘ§ ΝΉΥαΜ·ΓΔ ΦΉΜυΜ·ΦΑΖΚΥΊΜ·Β»Ζ≠“κΚσ–ό ΈΘέ13ΘίΘ§ “‘ΦΑΚ§Ζ«Χλ»ΜΑ±ΜυΥαΘέ14ΘίΒ»ΙΠΡήΒΡΒΑΑΉ÷ .
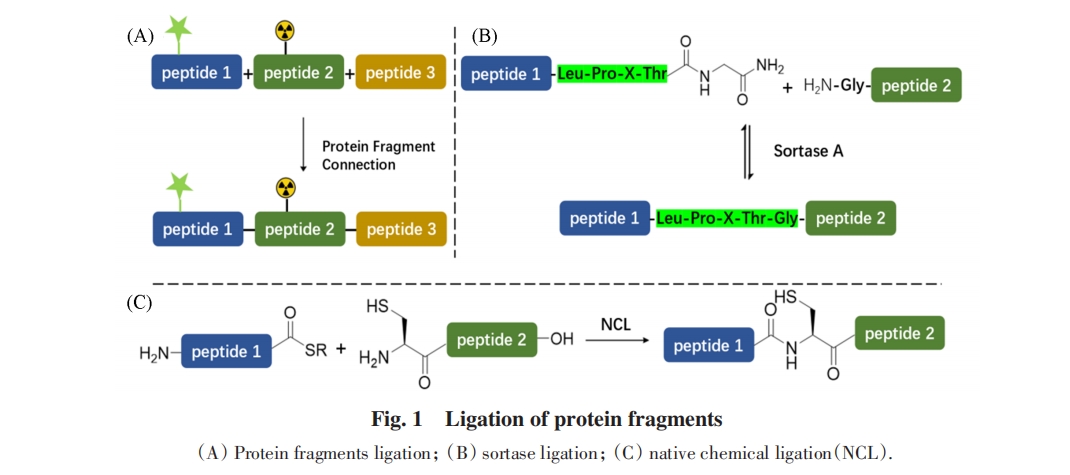
ΡΩ«ΑΘ§ ”Π”ΟΉνΙψΖΚΒΡΜ·―ßΝ§Ϋ”ΦΦ θ «Ή‘»ΜΜ·―ßΝ§Ϋ”Θ®NCLΘ©. ΗΟΦΦ θ”…÷ΞΦ”Ηγ¥σ―ßΒΡKentΒ»Θέ15Θί”Ύ1994ΡξΧα≥ωΘ§ Ά®Ιΐ“ΜΗωΕύκΡΤ§ΕΈΒΡC-ΕΥΝρθΞ”κΝμ“ΜΗωN-ΕΥCysΕύκΡΤ§ΕΈΒΡ―Γ‘ώ–‘Μ·―ßΖ¥”ΠΘ§ ‘ΎΝ§Ϋ”ΈΜΒψ¥Π–Έ≥…Χλ»ΜθΘΑΖΦϋ. ‘γΤΎΘ§ C-ΕΥΝρθΞΒΡΚœ≥… ήΒΫΈ»Ε®–‘≤νΒΡάß»≈ΘΜΫϋΡξά¥Ά®ΙΐθΘκ¬―θΜ·ΒΡC-ΕΥΝρθΞ‘≠ΈΜΚœ≥…ΦΑΝ§Ϋ”≤Ώ¬‘ΦΪ¥σΒΊ¥ΌΫχΝΥ’β“ΜΖΫΖ®ΒΡ”Π”ΟΘέ16~20Θί. »ΜΕχΘ§ ”…”ΎNCLΖ¥”Π“άΩΩΥφΜζ≈ωΉ≤Θ§ ΤδΝ§Ϋ”–߬ ήΒΉΈο≈®Ε»ΒΡ÷Τ‘ΦΘ§ NCLΆ®≥Θ–η“Σmmol/LΦΕ±πΒΡΒΉΈο≈®Ε». Β±ΒΉΈο≈®Ε»ΒΆ”Ύ1 mmol/L ±Ν§Ϋ”–߬ ΒΆΘ§ ΙΒΟNCL‘Ύ÷§ΒΑΑΉΓΔ ΡΛΒΑΑΉΒ»Ρ―»ήΒΑΑΉΤ§ΕΈΝ§Ϋ” ±≤ζ…ζάßΡ―. ΈΣ‘ωΦ”NCL‘ΎΒΆ≈®Ε»ΧθΦΰœ¬ΒΡΖ¥”Π–ß¬ Θ§ ―–ΨΩ’ΏΟ«Χα≥ωΝΥΕύ÷÷ΫβΨωΖΫΑΗΘ§ ÷ς“Σ”–ΘΚœρΖ¥”ΠΧεœΒΦ”»κ”–Μζ»ήΦΝΜρ’Ώ»ΞΙΗΦΝ“‘‘ωΦ”Τ§ΕΈ»ήΫβΕ»Θέ21ΘίΘΜ‘Ύ÷ςΝ¥…œ“ΐ»κΩ…“Τ≥ΐΒΡ‘ω»ή–ό Έ“‘¥ρΤΤ«βΦϋΆχ¬γΓΔ œς»θΨέΦ·«ψœρΘέ22Θ§23ΘίΘΜΆ®ΙΐDNA-DNAΓΔ ΒΑΑΉ-ΒΑΑΉ/–ΓΖ÷Ή”Β»«ΩœύΜΞΉς”ΟΡΘΑεΗ®÷ζΤ§ΕΈΝ§Ϋ”Θ§ ΙΤ§ΕΈΝ§Ϋ”‘ΎΫϋΥΤΖ÷Ή”ΡΎΖ¥”ΠΒΡΧθΦΰœ¬Ϋχ––Θέ24ΘίΒ». ¥ΥΆβΘ§ Ι”ΟΜν–‘ΗϋΗΏΒΡΈχθΞ¥ζΧφΝρθΞΉςΈΣθΘΜυΙ©ΧεΒΡΖΫΑΗ“≤Ρή ΙΒΟΖ¥”Π‘ΎΗϋΒΆ≈®Ε»œ¬Ϋχ––Θέ25Θί. Υδ»Μ’β–©≤Ώ¬‘”–÷ζ”ΎΚœ≥…ΨΏ”–Χτ’Ϋ–‘ΒΡΡΩ±ξΘ§ ΒΪ“≤¥φ‘Ύ‘ωΦ”Α≤ΉΑΚΆ“Τ≥ΐ±ξ«©ΜρΡΘΑεΒΡΕνΆβ≤ΌΉςΓΔ ±ξ«©ΉνΦ―Α≤ΉΑΈΜ÷ΟΡ―“‘‘Λ≤βΦΑΈχθΞΒΡΈ»Ε®–‘ΫœΒΆΒ»ΙΧ”–Έ ΧβΘ§ ΒΦ÷¬’β–©ΖΫΖ®‘ΎΈ»ΫΓ–‘ΚΆ≤ΌΉς±ψΫί–‘ΖΫΟφ¥φ‘Ύ≤ΜΉψ.
Νμ“ΜΖΫΟφΘ§ Μ·―ßΟΗΖ®“ρΤδΡήΙΜ‘ΎΒΆ≈®Ε»œ¬ΗΏ–ßΖ¥”ΠΘ§ ‘ΎΒΑΑΉ÷ Κœ≥…÷–“≤±ΜΙψΖΚ”Π”ΟΘέ26~28Θί. ΒΪ’βάύΝ§Ϋ”Ζ¥”ΠΆ®≥Θ–η“Σ‘ΎΒΉΈο÷–“ΐ»κΟΗΧΊ“λ–‘ Ε±π–ρΝ–Θ§ ’β–© Ε±π–ρΝ–‘ΎΖ¥”ΠΚσΆυΆυΜα±ΘΝτ‘Ύ≤ζΈοΒΑΑΉΒΡ–ρΝ–÷–Θ§ Ντœ¬–ρΝ–ΓΑΑΧΚέΓ±Θ§ »γSortase AΟΗΒΡ≤–Ντ–ρΝ–LPXTGΘέ29Θί. Υδ»ΜΩ…“‘Ά®Ιΐ―Γ‘ώΡΩ±ξΒΑΑΉ÷ ΙΧ”–¥φ‘ΎΒΡΟΗΒΡ Ε±π–ρΝ–ΉςΈΣΝ§Ϋ”ΈΜΒψΘ§ ΒΪ’β–©«ιΩω «ΦΪΤδ≈Φ»ΜΒΡΘΜΆ®ΙΐΗΏΆ®ΝΩ…Η―ΓΦΦ θΕ‘Sortase AΫχ–––ρΝ–ΫχΜ·Θέ30~33ΘίΘ§ “‘ΦΑ Ι”Ο‘Λœ»÷Τ±Η≥…ΝρθΞΒΡΒΑΑΉΤ§ΕΈΉςΈΣΝ§Ϋ”ΒΉΈοΘέ34ΘίΒ»≤Ώ¬‘Ω…“‘Φθ…ΌSortase AΟΗΒΡ–ρΝ–“άάΒ–‘Θ§ ≤ΔΦθ…ΌΖ¥”ΠΒΡΩ…Ρφ–‘Θέ35ΘίΘ§ ΒΪ“Σ Βœ÷Άξ»ΪΈόΚέΒΡΤ§ΕΈΝ§Ϋ”»‘»ΜάßΡ―.
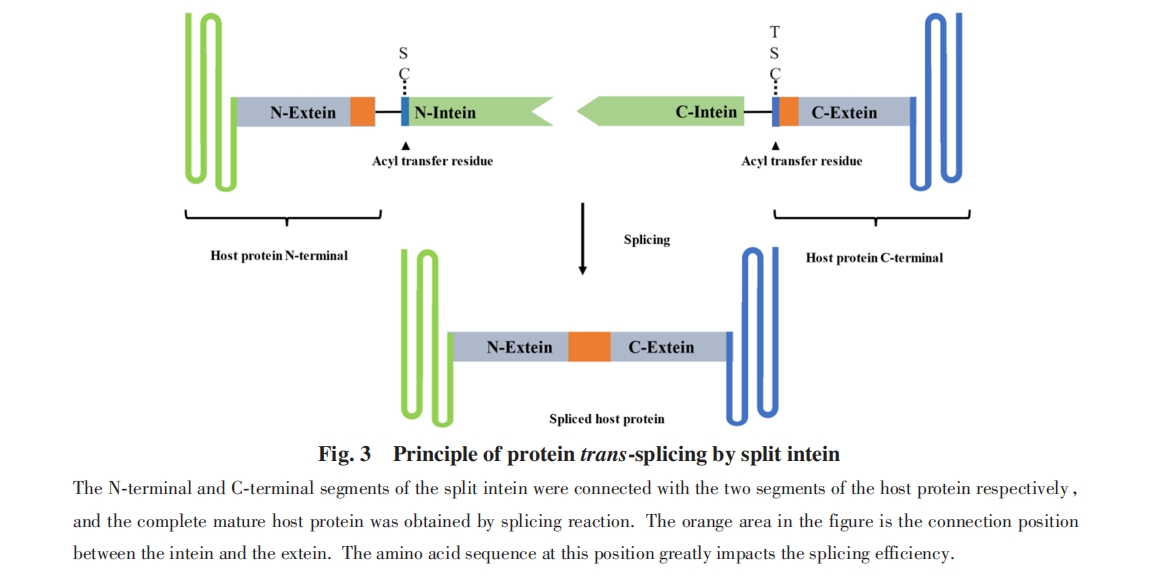
“ρ¥ΥΘ§ ―Α’““Μ÷÷ΡήΙΜΗΏ–ßΝ§Ϋ”ΒΑΑΉΤ§ΕΈ«“Ν§Ϋ”ΚσΈόΚέΒΡΒΑΑΉΝ§Ϋ”ΙΛΨΏΘ§ ≥…ΈΣ―–ΨΩ’ΏΙΊΉΔΒΡΖΫœρ. NCLΖ¥”ΠΚΆSortase AΟΗΝ§Ϋ”Ιΐ≥Χ÷–ΕΦΨ≠άζΝΥΝρθΞΚΆθΘΜυS-N«®“ΤΙΐ≥ΧΘΜ‘Ύ…ζΟϋΫχΜ·Ιΐ≥Χ÷–Θ§ ΖΚΥΊΜ·ΟΗΦΕΝΣΖ¥”ΠΒ»Ιΐ≥Χ÷–“≤Ιέ≤λΒΫΝρθΞ÷–ΦδΧεΚΆθΘΜυS-N«®“ΤΙΐ≥ΧΘέ36ΘίΘ§ ”…¥ΥΩ…Φϊ’β «…ζ≥…–¬ΒΡκΡΦϋΒΡ“Μ÷÷”––ßΆΨΨΕ. ¬ Β…œΘ§ »ΥΟ«“―Ψ≠ΖΔœ÷ΝΥ“ΜάύΡήΙΜΆ®ΙΐΝρθΞ÷–ΦδΧεΚΆθΘΜυ«®“Τ¬ΖΨΕ Βœ÷Ε‘ΒΑΑΉ÷ Τ§ΕΈΒΡΗΏ–ßΈόΚέΝ§Ϋ”ΒΡΒΑΑΉΘΚΡΎΚ§κΡΘ®InteinΘ©. ΡΎΚ§κΡΩ…“‘Ζ÷ΈΣΥ≥ ΫΡΎΚ§κΡΘ®Cis-inteinΘ©”κΖ÷Ν―ΡΎΚ§κΡΘ®Split inteinΘ©ΝΫ¥σάύ. Τδ÷–Θ§ Ζ÷Ν―ΡΎΚ§κΡΩ…“‘”κ–η“ΣΫχ––Ν§Ϋ”ΒΡΒΑΑΉΤ§ΕΈΖ÷±πœύΝ§Θ§ “―Ψ≠‘Ύ¥σΝΩΒΑΑΉ÷ Ν§Ϋ””κΚœ≥…÷–’Ιœ÷≥ω”Π”ΟΦέ÷ΒΘ§ ήΒΫΝΥ―–ΨΩ’ΏΒΡΙψΖΚΙΊΉΔ. ±ΨΈΡ÷ς“ΣΫι…ήΖ÷Ν―ΡΎΚ§κΡΉςΈΣ“Μ÷÷ΒΑΑΉ÷ Κœ≥…ΙΛΨΏΘ§ ΫαΚœΖ÷Ν―ΡΎΚ§κΡ‘ΎΒΑΑΉ÷ Κœ≥…–ό ΈΝλ”ρΒΡ”Π”ΟΑΗάΐΘ§ Ζ÷ΈωΝΥΖ÷Ν―ΡΎΚ§κΡœύ±»ΤδΥϋΖΫΖ®ΒΡ”≈ ΤΘ§ ΉήΫαΖ÷Ν―ΡΎΚ§κΡΒΡΖΔ’Ιœ÷Ή¥Θ§ ≤Δ’ΙΆϊΝΥΤδΈ¥ά¥ΒΡΖΔ’Ι«ΑΨΑ.
1 Ζ÷Ν―ΡΎΚ§κΡΒΡΖΔœ÷
1.1ΓΓΡΎΚ§κΡΒΡΖΔœ÷
1990ΡξΘ§ StevensΒ»ΖΔœ÷ΡπΨΤΫΆΡΗΘ®Saccharomy cescerevisiaeΘ©TFP1Μυ“ρΒΡΖ≠“κ≤ζΈοΜα‘ΎΖ≠“κΚσ±Μ«–Ην≥…2ΗωΒΑΑΉΘ§ ÷–―κΒΡ–ρΝ–Φτ«–≤ζ…ζΖ÷Ή”ΝΩΈΣ50000ΒΡΒΑΑΉΘ§ ΝΫΕΥΒΡΤ§ΕΈΤ¥Ϋ”–Έ≥…Ζ÷Ή”ΝΩΈΣ69000ΒΡΒΑΑΉ. ¥ΥΚσ―–ΨΩ’Ώ‘ΎΗϋΕύΈο÷÷÷–ΖΔœ÷ΝΥάύΥΤΒΡœ÷œσΘέ37Θ§38Θί. Κσ–χ―–ΨΩΖΔœ÷Θ§ ’β÷÷≤ε»κΥό÷ςΒΑΑΉ–ρΝ–ΒΡΒΑΑΉΩ…“‘¥ΏΜ·Ή‘…μ¥”Υό÷ςΒΑΑΉ÷–ΕœΝ―Θ§ Ά§ ± ΙΝΫ≤ύΒΡΥό÷ςΒΑΑΉΤ§ΕΈΆ®ΙΐκΡΦϋΝ§Ϋ”Τπά¥–Έ≥…≥… λΒΑΑΉΘέ39Θί. ’β÷÷ΨΏ”–ΦτΫ”ΝΫ≤ύΒΑΑΉΤ§ΕΈΡήΝΠΒΡΒΑΑΉ±ΜΟϋΟϊΈΣΡΎΚ§κΡΘ®InteinΘ©Θ§ ΕχΝΫΕΥ±ΜΦτΫ”ΒΡΒΑΑΉΤ§ΕΈ±ΜΟϋΟϊΈΣΆβœ‘κΡΘ®ExteinΘ©Θ®ΆΦ2Θ©.

1.2ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΖΔœ÷
Ζ÷Ν―ΡΎΚ§κΡ «Ζ÷Ν―ΈΣ2ΗωΕάΝΔ¥φ‘ΎΒΡ«ΑΧεΤ§ΕΈΒΡΡΎΚ§κΡΘ§ ΟΩΗωΤ§ΕΈΖ÷±π”κ1ΗωΆβœ‘κΡ»ΎΚœΘ®ΆΦ3Θ©. 2Ηω«ΑΧεΤ§ΕΈΖ«Ι≤ΦέΫαΚœ≤ΔΉιΉΑ≥…Βδ–ΆΒΡΡΎΚ§κΡΫαΙΙΘ§ ¥”ΕχΜ÷Η¥Ζ÷Ν―ΡΎΚ§κΡΒΡΦτΫ”Μν–‘Θ§ ¥ΏΜ·ΝΫ≤ύΒΡΆβœ‘κΡΖΔ…ζΒΑΑΉ÷ Ζ¥ ΫΦτΫ”Θ®PTSΘ©. ΤδΩ…“‘ά¥‘¥”ΎΧλ»ΜΝ§–χΡΎΚ§κΡΒΡ»ΥΙΛΖ÷Ν―Θ§ “≤‘ΎΉ‘»ΜΫχΜ·÷–¥φ‘Ύ.
1969ΡξΘ§ AnfinsenΒ»Θέ40ΘίΖΔœ÷‘ΎΧΊΕ®ΒΡΧθΦΰœ¬Θ§ Ζ÷Ν―≥…2ΗωΤ§ΕΈΒΡΟΗΡήΙΜΖ«Ι≤ΦέΫαΚœ≤ΔΜ÷Η¥Άξ’ϊΟΗΒΡ¥ΏΜ·ΙΠΡή. 1998ΡξΘ§ PerlerΒ»Θέ41ΘίΫΪ’β“ΜΥΦœκ‘Υ”ΟΒΫΡΎΚ§κΡΒΡ…ηΦΤΗΡ‘λ÷–. ΥϊΟ«ΫΪPsp Pol-1ΡΎΚ§κΡ÷–ΒΡ“ΜΗωκΡΦϋ«–ΕœΘ§ ΫΪΤδΖ÷Ν―ΈΣN-ΕΥΤ§ΕΈΘ®IntNΘ©ΚΆC-ΕΥΤ§ΕΈΘ®IntCΘ©Θ§ ΫΪ2ΗωΤ§ΕΈΖ÷±π±μ¥ο¥ΩΜ·Κσ‘ΎΧεΆβΙ≤Η¥–‘Θ§ ΫαΙϊ±μΟς2ΗωΤ§ΕΈΆ®ΙΐΖ«Ι≤ΦέΉς”Ο–Έ≥…Η¥ΚœΈο≤ΔΜ÷Η¥ΦτΫ”Μν–‘.
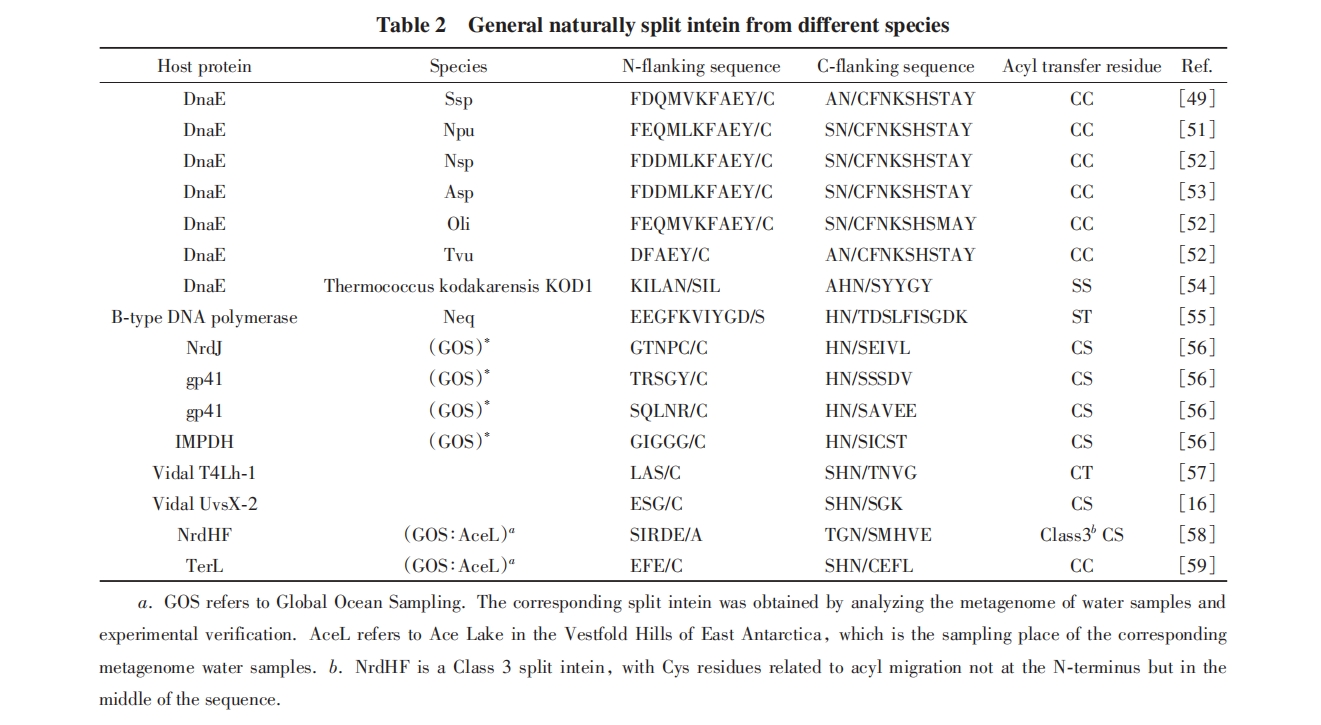
Ά§ΡξΘ§ PaulusΒ»Θέ42ΘίΫΪMtu RecAΡΎΚ§κΡΖ÷Ν―ΈΣN-ΕΥΚΆC-ΕΥ2ΗωΤ§ΕΈΘ§ ≤ΔΗςΉ‘»ΎΚœΒΫœύ”ΠΒΡΆβœ‘κΡ…œΘ§ Ζ÷±π±μ¥ο¥ΩΜ·ΚσΙ≤Η¥–‘Θ§ 2ΗωΤ§ΕΈΫαΚœ–Έ≥…ΝΥΨΏ”–ΦτΫ”ΙΠΡήΒΡΗ¥ΚœΈο. ΧΫΨΩΝΥΦτΫ”Ζ¥”ΠΙΐ≥ΧΕ‘―θΜ·ΜΙ‘≠ΜΖΨ≥ΚΆpH÷ΒΒΡ“άάΒ–‘Θ§ ≤Δ÷Η≥ω’β÷÷ΦτΫ”œΒΆ≥ «“Μ÷÷”–«±ΝΠΒΡΒΑΑΉ÷ Ν§Ϋ”ΙΛΨΏ. ¥ΥΚσΘ§ ―–ΨΩ’ΏΕ‘Χλ»ΜΝ§–χΡΎΚ§κΡΫχ––ΝΥ¥σΝΩΒΡ»ΥΙΛΕœΝ―≥Δ ‘Θ§ ΖΔ’Ι≥ωΕύ÷÷»ΥΙΛΖ÷Ν―ΡΎΚ§κΡΘ®±μ1Θ©.
1998ΡξΘ§ LiuΒ»Θέ49Θί‘ΎSsp DnaEΘ®άΕ‘εΒΡDNAΨέΚœΟΗΔσΒΡ¥ΏΜ·―«ΜυAΘ©ΒΑΑΉ÷–ΦχΕ®≥ω ΉΗωΧλ»ΜΒΡΖ÷Ν―ΡΎΚ§κΡ. DnaEΒΡN-ΕΥΚΆC-ΕΥ≤ΩΖ÷”…2ΗωΒΞΕάΒΡΜυ“ρDnaE-NΚΆDnaE-CΖ÷±π±ύ¬κ. ‘Ύ¥σ≥ΠΗΥΨζ÷–≤β ‘ ±Θ§ 2ΗωΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΆ®ΙΐΖ«Ι≤ΦέΉς”ΟΫαΚœ–Έ≥…ΡΎΚ§κΡΗ¥ΚœΈο. ΗΟΗ¥ΚœΈο≤ΜΫωΨΏ”–άύΥΤΡΎΚ§κΡΒΡ–ρΝ–ΧΊ’ςΘ§ Εχ«“±μœ÷≥ωΒΑΑΉ÷ Ζ¥ ΫΦτΫ”Μν–‘Θ§ ΦτΫ”ΚσΒΟΒΫΆξ’ϊΒΡDNAΨέΚœΟΗΔσ―«ΜυA. ’β“ΜΖΔœ÷±μΟςΘ§ Ζ÷Ν―ΡΎΚ§κΡ «Χλ»Μ¥φ‘ΎΒΡ. ”κ»ΥΙΛΖ÷Ν―ΡΎΚ§κΡœύ±»Θ§ Χλ»ΜΖ÷Ν―ΡΎΚ§κΡΨΏ”–Ω…‘ΎΖ«±δ–‘ΧθΦΰœ¬ΉιΉΑΦΑΦτΫ”–߬ ΗϋΗΏΒ»”≈ Τ.
2009ΡξΘ§ MootzΒ»Θέ50ΘίΖΔœ÷ΝΥΝμ“Μ÷÷–¬ΒΡΧλ»ΜΖ÷Ν―ΡΎΚ§κΡNpu DnaE. Npu DnaEΨΏ”–ΦΪΗΏΒΡΦτΫ”Ζ¥”ΠΜν–‘Θ§ ‘Ύ37 Γφœ¬Ζ¥ ΫΦτΫ”Ζ¥”ΠΒΡΑκΥΞΤΎΘ®Φ¥Ζ¥”ΠΈοΒΡ≈®Ε»œϊΚΡ“ΜΑκ ±Υυ–ηΒΡ ±Φδt1/2Θ§ sΘ©ΫωΈΣ63 sΘ§ ΦτΫ”Ζ¥”ΠΥΌ¬ ≥Θ ΐkΘ®s-1Θ©>3.5ΓΝ10-3 s-1. ¥ΥΚσΘ§ ―–ΨΩ’ΏΟ«Ά®ΙΐΜυ“ρ ΐΨί±»Ε‘ΚΆ Β―ι―ι÷ΛΒ» ÷ΕΈΘ§ ΖΔœ÷ΝΥΗϋΕύά¥Ή‘≤ΜΆ§Έο÷÷ΒΡΧλ»ΜΖ÷Ν―ΡΎΚ§κΡΘ®±μ2Θ©Θ§ Ϋχ“Μ≤Ϋά©¥σΝΥΖ÷Ν―ΡΎΚ§κΡΉςΈΣΒΑΑΉ÷ Ν§Ϋ”ΙΛΨΏΒΡΩ…–––‘”κ”Π”Ο«ΑΨΑ.
1.3ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΖ÷άύ
Ζ÷Ν―ΡΎΚ§κΡΩ…“‘ΗυΨίΤδΦΡ÷ςΜυ“ρΙιάύΈΣΨΏ”–“≈¥ΪΆ§‘¥ΙΊœΒΒΡΦ“Ήεάύ. ¥ΥΆβΘ§ ≤ΜΆ§ΒΡΖ÷Ν―ΡΎΚ§κΡ‘ΎΖ¥”ΠΙΐ≥Χ÷–÷–ΦδΧε¥φ‘Ύ≤ν“λΘ§ “≤Ω…“‘Ψί¥ΥΕ‘Ζ÷Ν―ΡΎΚ§κΡΫχ––άύ±πΜ°Ζ÷. ‘ΎΫαΙΙ…œΘ§ ΡΎΚ§κΡ“ΜΑψ≥ œ÷ΕύΗωΠ¬Τ§≤ψΒΡΫαΙΙΘ§ Ζ÷Ν―ΒΡΈΜΒψ“ΜΑψ‘ΎΠ¬Τ§≤ψ÷°ΦδΒΡLoop«χΘ§ ¥”≤ΜΆ§«χ”ρΕœΝ―ΒΡΡΎΚ§κΡΨΏ”–≤ΜΆ§ΒΡΧΊ–‘Θ§ ΡήΙΜΖαΗΜΖ÷Ν―ΡΎΚ§κΡΒΡ”Π”Ο≥ΓΨΑΘ§ “ρ¥Υ―–ΨΩ’Ώ“≤ΗυΨίΖ÷Ν―ΡΎΚ§κΡΒΡΖ÷Ν―ΈΜΒψΕ‘ΤδΫχ––ΙιΡ….
‘Φ70%ΒΡΡΎΚ§κΡΒΡΥό÷ςΒΑΑΉ”κDNAΗ¥÷ΤΚΆDNA–όΗ¥”–ΙΊΘέ60ΘίΘ§ ≤Δ«“‘ΎΫχΜ·ΖΔ’ΙΙΐ≥Χ÷–±μœ÷≥ωΜυ“ρΥ°ΤΫ«®“ΤΒΡΧΊ’ςΘέ61Θί. “ρ¥ΥΖ÷Ν―ΡΎΚ§κΡΩ…“‘Α¥’’Τδά¥‘¥ΒΡΥό÷ςΜυ“ρΫχ––Ζ÷άύΘ§ ά¥‘¥”ΎΆ§άύΜυ“ρΒΡΖ÷Ν―ΡΎΚ§κΡΒΡ–ρΝ–Ά§‘¥–‘ΫœΗΏΘ§ ≤Δ«“ΨΏ”–άύΥΤΒΡΧΊ’ς. ά¥‘¥”Ύ≤ΜΆ§Έο÷÷ΒΡDna EΖ÷Ν―ΡΎΚ§κΡΕΦΡή‘ΎœύΆ§ΒΡΆβœ‘κΡ–ρΝ–Ϋχ––«–ΗνΘ§ ≤Δ«“―–ΨΩ“≤ΖΔœ÷ΤδΦδ¥φ‘ΎΫΜ≤φΖ¥”Π–‘Θ§ »γNpu DnaEΒΡIntNΤ§ΕΈΚΆSsp DnaEΒΡIntCΤ§ΕΈ“≤Ω…“‘Ζ«Ι≤ΦέΫαΚœ≤Δ–Έ≥…ΨΏ”–ΦτΫ”Ζ¥”ΠΜν–‘ΒΡΗ¥ΚœΧεΘέ62Θί. ―–ΨΩ’ΏΕ‘”Ύ‘¥”ΎDnaEΘ§ GyrBΚΆDnaBΒ»Φ“ΉεΒΡΖ÷Ν―ΡΎΚ§κΡ“―Ψ≠Ϋχ––ΝΥœΒΆ≥ΒΡ―–ΨΩΘ§ ≤Δ«“ΩΣΖΔΝΥΕύ÷÷”Π”Ο.
Ζ÷Ν―ΡΎΚ§κΡ¥ΏΜ·ΒΡΦτΫ”Ζ¥”ΠΙΐ≥Χ…φΦΑΕύ≤ΫΙΊΦϋΒΡθΘΜυΉΣ“ΤΖ¥”ΠΘ§ ΙΊΦϋΑ±ΜυΥα≤–Μυ‘Ύ’β–©θΘΜυΉΣ“ΤΙΐ≥Χ÷––Έ≥…ΝρθΞΜρ’Ώ―θθΞ÷–ΦδΧεΘ§ Ε‘Ζ¥”Π–‘÷ ≤ζ…ζœ‘÷χ”Αœλ. “ΜΑψΕχ―‘Θ§ ΝρθΞ÷–ΦδΧεΈ»Ε®–‘Ϋœ≤νΘ§ ‘ΎΧεΆβ”Π”Ο÷–Ψ≠≥ΘΙέ≤λΒΫ÷–ΦδΧεΝρθΞΦϋΥ°ΫβΒΦ÷¬ΒΡΗ±≤ζΈοΘ§ Εχ≤…”Ο―θθΞ÷–ΦδΧεΒΡΖ÷Ν―ΡΎΚ§κΡ‘ρΖ¥”ΠΥΌ¬ Ϋœ¬ΐ. “ρ¥Υ―–ΨΩ’ΏΗυΨίΖ÷Ν―ΡΎΚ§κΡ1ΈΜΚΆCΕΥΆβœ‘κΡ+1ΈΜΒΡΑ±ΜυΥαΘ§ ΫΪΤδΖ÷ΈΣCCΚΆCSΘ®TΘ©ΝΫάύ. CCάύΖ÷Ν―ΡΎΚ§κΡΘ®»γSsp DnaEΦΑSce VMAΒ»Θ©1ΈΜΚΆCΕΥΆβœ‘κΡ+1ΈΜΒΡΑ±ΜυΥαΨυΈΣΑκκΉΑ±ΥαΘ§ Ζ¥”ΠΨ≠άζ2ΗωΝρθΞ÷–ΦδΧεΙΐ≥Χ. CSΘ®TΘ©άύΖ÷Ν―ΡΎΚ§κΡΘ®»γSsp DnaBΘ§ gp41Β»Θ©1ΈΜΈΣΑκκΉΑ±ΥαΘ§ CΕΥΆβœ‘κΡ+1ΈΜΈΣΥΩΑ±ΥαΜρΥ’Α±ΥαΘ§ Ζ¥”Π÷–Ψ≠άζ1ΗωΝρθΞ÷–ΦδΧεΚΆ1Ηω―θθΞ÷–ΦδΧεΙΐ≥Χ. ≥ΐ¥ΥΝΫ¥σάύΆβΘ§ ΜΙ”–…Ό ΐΖ÷Ν―ΡΎΚ§κΡ‘ΎΖ¥”Π÷–Ψ≠άζ2Ηω―θθΞ÷–ΦδΧεΙΐ≥ΧΘ§ »γNeq DNA polymeraseΖ÷Ν―ΡΎΚ§κΡΘέ55Θί.
Ζ÷Ν―ΡΎΚ§κΡ“ΜΑψΖ÷ΈΣ2ΗωΤ§ΕΈΘ§ 2ΗωΤ§ΕΈΖ«Ι≤ΦέΫαΚœΚσ‘ΎΫαΙΙΚΆΙΠΡή…œΫ”Ϋϋ”ΎΝ§–χΒΡΡΎΚ§κΡ. Έό¬έ «»ΥΙΛΖ÷Ν―ΒΡΡΎΚ§κΡΜΙ «Χλ»Μά¥‘¥ΒΡΖ÷Ν―ΡΎΚ§κΡΘ§ ΤδΖ÷Ν―ΈΜΒψΨυΈΜ”ΎΫαΙΙ…œΒΡΜΖ«χΘ®LoopΘ©Θ§ ¥”Εχ±ήΟβ”ΑœλΫαΚœΚΆΦτΫ”Ζ¥”ΠΜν–‘. ΗυΨίΖ÷Ν―ΈΜΒψ≤ΜΆ§Θ§ ΡΎΚ§κΡΩ…“‘±ΜΖ÷ΈΣS0Θ§ S1ΚΆS11ΡΎΚ§κΡΘέ63Θί. S0ΡΎΚ§κΡ¥”‘≠ ΦΡΎΚ§κΡ÷–Ιι≥≤ΚΥΥαΡΎ«–«χ”ρΈΜΒψ«–ΗνΘ§ »γSsp DnaEΚΆNpu DnaEΒ». S1ΡΎΚ§κΡ¥”Ϋ”ΫϋNΕΥΒΡΈΜ÷Ο«–ΗνΘ§ »γSsp DnaBΚΆM86 DnaBΒ». S11ΡΎΚ§κΡ¥”Ϋ”ΫϋCΕΥΒΡΈΜ÷Ο«–ΗνΘ§ »γSsp GyrB S11ΚΆSsp DnaB S11Β». ¥ΥΆβΘ§ LiuΒ»Θέ63ΘίΫΪDnaB¥”S1ΚΆS0ΈΜΒψΖ÷ΈΣ3ΕΈΘ§ “ά»Μ±ΘΝτΫαΚœΦΑΦτΫ”ΙΠΡή.
2 Ζ÷Ν―ΡΎΚ§κΡΒΡΖ¥”Π‘≠άμ
2.1ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΫαΙΙ
Ζ÷Ν―ΡΎΚ§κΡΩ…“‘Ω¥Ής «Άξ’ϊΡΎΚ§κΡΒΡΖ÷Ν―–Έ ΫΘ§ ΤδΗ¥ΚœΈοΒΡΫαΙΙ”κΆξ’ϊΡΎΚ§κΡœύΥΤΘέ64ΘίΘ§ «Έ±C2Ε‘≥Τ–‘ΒΡ¬μΧψ–ΈΒΑΑΉΘ§ ÷ς“Σ”…Π¬’έΒΰΉι≥…. ΝΫΉιΖ¥ΤΫ––ΒΡΠ¬’έΒΰ≥…¬μΧψΒΡΤΫΟφΘ§ ΤΫΟφ…œœ¬Ης”–“ΜΉι2ΗωΠ¬’έΒΰΚΆ“ΜΧθΠΝ¬ί–ΐ–Έ≥…ΒΡΫαΙΙ”ρΘ®Π¬Π¬ΠΝ motifΘ©Θ§ Τδ÷–“Μ≤ύΒΡΠ¬’έΒΰ―”…λ≤Δ¥Ι÷±¥©Ιΐ¬μΧψ÷–―κΒΡloopΘ§ ”κNΕΥΆβœ‘κΡœύΝ§. ¬μΧψ÷–―κΝμ“ΜΕ‘¥Ι÷±”ΎΤΫΟφΒΡΠ¬’έΒΰ”κCΕΥΆβœ‘κΡœύΝ§. ΝΫΕΈΆβœ‘κΡΈΜ”ΎΤΫΟφΒΡΆ§“Μ≤ύΘέΆΦ4Θ®AΘ©ΚΆΘ®CΘ©ΘίΘέ65Θί.
Ζ÷Ν―ΡΎΚ§κΡ”–Εύ÷÷Ω…ΡήΒΡΖ÷Ν―ΖΫ ΫΘ§ Εύ ΐΖ÷Ν―ΡΎΚ§κΡΖ÷Ν―≥…“Μ¥σ“Μ–Γ2ΗωΤ§ΕΈΘ§ Ζ÷Ν―ΈΜΒψΩ…Ρή‘ΎΩΩΫϋCΕΥΒΡ“Μ≤ύΘ§ “≤Ω…Ρή‘ΎΩΩΫϋNΕΥΒΡ“Μ≤ύ. Τδ÷–Θ§ ΩΩΫϋCΕΥΒΡΖ÷Ν―±Μ≥ΤΈΣΨ≠ΒδΘ®TypicalΘ©Ζ÷Ν―Θ§ Τδ–ΓΤ§ΕΈ“ΜΑψΑϋά®÷–―κ¥Ι÷±ΒΡ“ΜΉιΠ¬’έΒΰΚΆ»ΤΤΫΟφΠ¬’έΒΰΒΡ“Μ≤ΩΖ÷. ΩΩΫϋNΕΥΒΡΖ÷Ν―±Μ≥ΤΈΣΖ«Ψ≠ΒδΘ®AtypicalΘ©Ζ÷Ν―Θ§ Τδ–ΓΤ§ΕΈ“ΜΑψΑϋά®÷–―κ¥Ι÷±ΒΡloopΚΆΤΫΟφ“Μ≤ύΠ¬Π¬ΠΝ motifΒΡ“Μ≤ΩΖ÷. ¥ΥΆβΘ§ LiuΒ»Θέ63ΘίΆ®Ιΐ»ΥΙΛ…Η―ΓSsp DnaBΩ…ΡήΒΡΖ÷Ν―ΈΜΒψΘ§ Βœ÷ΝΥ3ΕΈΖ÷Ν―ΡΎΚ§κΡΒΡΦτ«–Ζ¥”Π. “―Ψ≠‘ΎNpu DnaE…œ Βœ÷άύΥΤΒΡ3Τ§ΕΈΖ÷Ν―ΡΎΚ§κΡΦτ«–Ζ¥”ΠΘ§ ΥΒΟςΆ®ΙΐΗΡ‘λΒΟΒΫΕύ”Ύ2Τ§ΕΈΒΡΖ÷Ν―ΡΎΚ§κΡ «Ω…ΡήΒΡΘέΆΦ4Θ®BΘ©ΚΆΘ®CΘ©Θί.
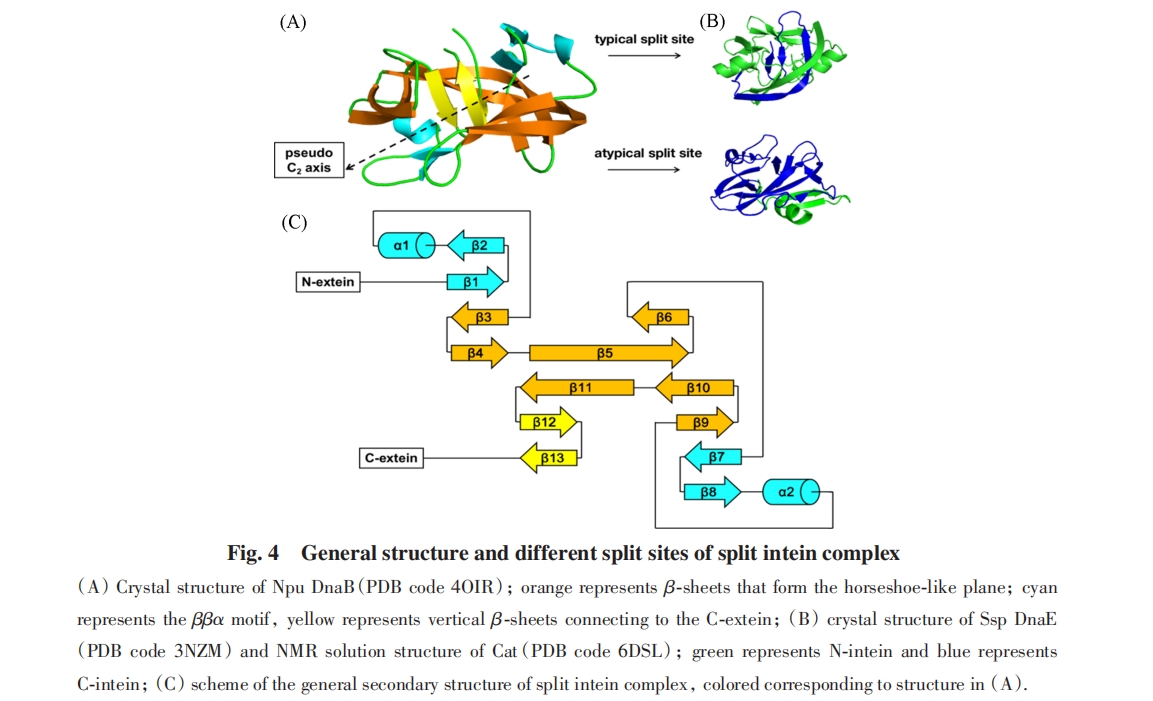
Ζ÷Ν―ΡΎΚ§κΡ–η“ΣΗςΉιΖ÷ΫαΚœΚσ≤≈Ρή’έΒΰ≥…’ΐ»ΖΒΡΙΙœσ. 2013ΡξΘ§ MuirΒ»Θέ66ΘίΆ®ΙΐΚΥ¥≈Ι≤’ώ≤®ΤΉΘ®NMRΘ©Ιέ≤βΝΥ»ή“Κ÷–Npu DnaEΖ÷Ν―ΡΎΚ§κΡΫαΚœ«ΑΚσΒΡΙΙœσΘ§ ΖΔœ÷Ζ÷Ν―ΡΎΚ§κΡΤ§ΕΈΫαΚœ«ΑΘ§ –ΓΤ§ΕΈ¥Π”ΎΈό–ρΉ¥Χ§Θ§ ¥σΤ§ΕΈ¥Π”ΎΙΙœσΕ·Χ§±δΜ·ΒΡΉ¥Χ§. ΝΫΤ§ΕΈΒΡœύΜΞΉς”Ο”’ΒΦ–ΓΤ§ΕΈ¥”Έό–ρΫαΙΙ’έΒΰΈΣΈ»Ε®ΙΙœσΘ§ ≤Δ”’ΒΦ¥σΤ§ΕΈ–Έ≥…ΗϋΦ”Έ»Ε®ΒΡΫαΙΙ. άύΥΤΒΡNMR–≈Κ≈ΧΊ’ς“≤‘ΎSsp DnaEΚΆCatΝΫ÷÷Ζ÷Ν―ΡΎΚ§κΡ…œ±Μ―ι÷ΛΘέ67Θ§68Θί. 2018ΡξΘ§ Rib®°Β»Θέ64ΘίΫβΈωΝΥNeq B-type DNA polymerase1Θ®Neq Pol1Θ© NΕΥ¥σΤ§ΕΈΚΆΝΫΤ§ΕΈΗ¥ΚœΈοΒΡΨßΧεΫαΙΙΘ§ ΖΔœ÷IntCΒΞΕά¥φ‘ΎΒΡΫαΙΙ”κ–Έ≥…Η¥ΚœΈοΚσΒΡΫαΙΙ”–Οςœ‘«χ±π. ’β–©ΫαΙϊΥΒΟςΖ÷Ν―ΡΎΚ§κΡΗςΉιΖ÷ΈόΖ®ΕάΝΔΈ»Ε®’έΒΰ≥…’ΐ»ΖΒΡΙΙœσ.
2.2ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΫαΚœΡΘ Ϋ
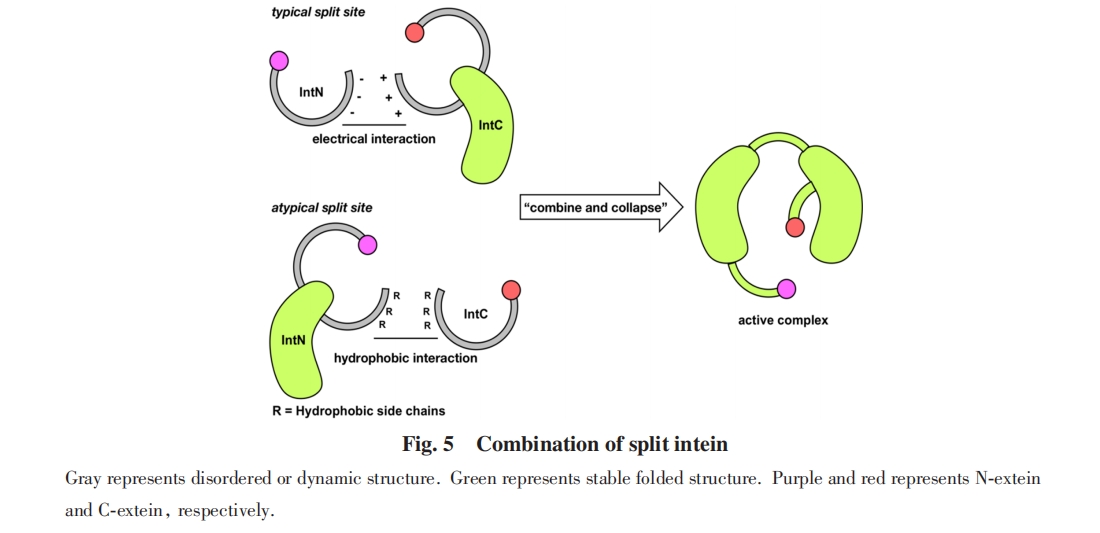
Ψ≤ΒγœύΜΞΉς”Ο±Μ»œΈΣ «Ζ÷Ν―ΡΎΚ§κΡΝΫΤ§ΕΈ Ε±πΚΆΫαΚœΒΡ÷Ί“ΣΕ·ΝΠ. 2007ΡξΘ§ MuirΒ»Θέ69ΘίΆ®Ιΐ–ρΝ–±»Ε‘ΖΔœ÷≤ΜΆ§Έο÷÷DnaEΖ÷Ν―ΡΎΚ§κΡœ‘ Ψ≥ω±Θ ΊΒΡΒγΚ…Ζ÷άκΧΊ’ςΘΚCΕΥ–ΓΤ§ΕΈΚ§”–Φν–‘≤–ΜυΦ·÷–Τ§ΕΈΘ§ NΕΥ¥σΤ§ΕΈΚ§”–Υα–‘≤–ΜυΦ·÷–Τ§ΕΈ. 2ΗωΒγ–‘œύΖ¥ΒΡΤ§ΕΈΫιΒΦΒΡΨ≤ΒγœύΜΞΉς”ΟΡΘ Ϋ‘ΎSsp DnaEΦΑ Neq Pol1ΒΡΨßΧεΫαΙΙ÷–ΕΦΒΟΒΫΝΥ―ι÷ΛΘέ52Θ§64Θί.
Ζ«Ψ≠ΒδΖ÷Ν―ΡΎΚ§κΡΝΫΤ§ΕΈΒΡΫαΚœΟφ”κΨ≠ΒδΖ÷Ν―ΡΎΚ§κΡ≤ΜΆ§. 2018ΡξΘ§ MuirΒ»Θέ67ΘίΆ®ΙΐNMRΫβΈωΝΥ“Μ÷÷Ζ«Ψ≠ΒδΖ÷Ν―ΡΎΚ§κΡCatΒΡΫαΙΙΘ§ ΖΔœ÷2ΗωΡΎΚ§κΡΤ§ΕΈΒΡœύΜΞΉς”ΟΫγΟφ÷ς“Σ « ηΥ°≤–ΜυΘ§ Εύ ΐ¥χΒγ≤–Μυ±©¬Ε‘Ύ»ήΦΝ÷–. …ζΜ· Β―ιΫαΙϊ“≤±μΟςΘ§ CatΝΫΗωΤ§ΕΈΒΡΫαΚœ«ΩΕ» ή»ή“ΚάκΉ”«ΩΕ»ΒΡ”ΑœλΚή–ΓΘ§ ±μΟς ηΥ°œύΜΞΉς”Ο‘Ύ“Μ–©Ζ«Ψ≠ΒδΖ÷Ν―ΡΎΚ§κΡΒΡΫαΚœ÷–ΤπΗϋ÷ς“ΣΒΡΉς”ΟΘ®ΆΦ5Θ©
2.3ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΦτ«–Μζάμ
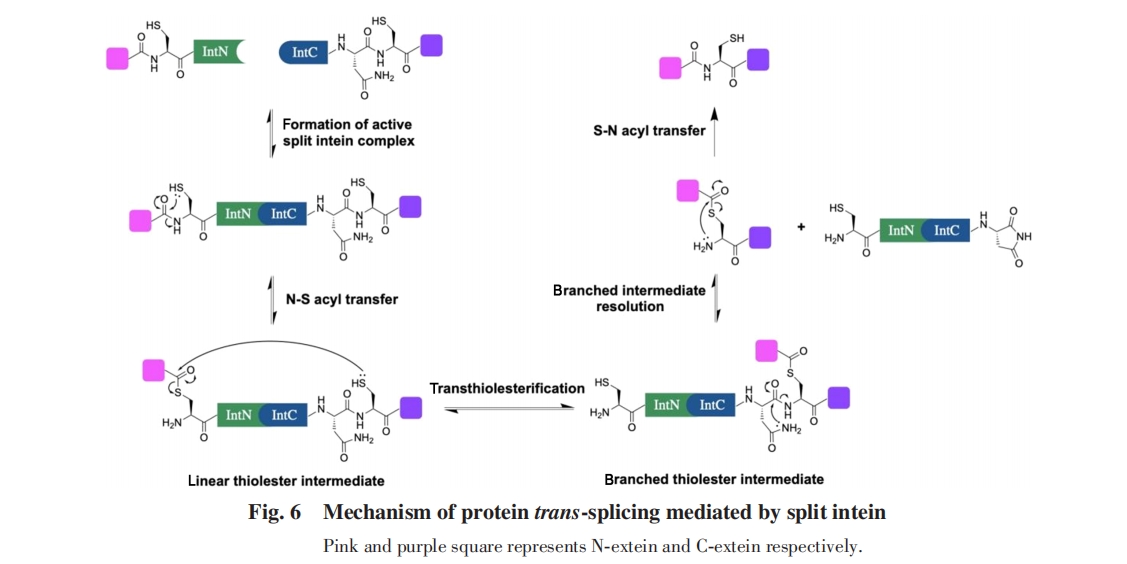
Ζ÷Ν―ΡΎΚ§κΡΒΡΦτΫ” «Ή‘ΖΔΙΐ≥ΧΘ§ ≤Μ–η“ΣΤδΥϋΒΡΜνΜ·Ρήά¥‘¥. ’ϊΗωΖ¥”ΠΑϋά®N-SθΘΜυ«®“ΤΓΔ ΝρθΞΫΜΜΜΓΔ ΧλΕ§θΘΑΖΜΖΜ·ΦΑS-NθΘΜυ«®“ΤΒ»4ΗωΙΐ≥ΧΘ®ΆΦ6Θ©.
ΒΎ“Μ≤ΫΘ§ ΡΎΚ§κΡN-ΕΥΒΡCys≤ύΝ¥έœΜυΫχΙΞ÷ςΝ¥θΘΑΖΦϋΘ§ ΖΔ…ζN-SθΘΜυ«®“ΤΘ§ ΙN-ΕΥΆβœ‘κΡΆ®ΙΐΝρθΞΦϋΝ§Ϋ”‘ΎΡΎΚ§κΡ…œ. ‘Ύ¥ΥΙΐ≥Χ÷–Θ§ Ζ÷Ν―ΡΎΚ§κΡ÷–÷Ν…Ό”–3ΗωΑ±ΜυΥαΤπΒΫ¥ΏΜ·Ής”ΟΘ§ Αϋά®”κέœΜυœύΜΞΉς”ΟΒΡF-Block Asp“‘ΦΑ”κθΘΑΖΦϋœύΜΞΉς”ΟΒΡB-Block HisΚΆB-Block Thr. F-Block AspΤπΒΫΦν¥ΏΜ·Ής”ΟΘ§ ¥ΌΫχCys≤ύΝ¥έœΜυΒΡ»Ξ÷ Ή”Μ·Θ§ ‘ω«ΩΝΥέœΜυ«ΉΚΥ–‘Θ§ ΒψΆΜ±δ Β―ι±μΟςΗΟΑ±ΜυΥαΕ‘N-SθΘΜυ«®“Τ «±Ί“ΣΒΡΘέ70Θί. …œ ωΜζάμ”κNMRΙέ≤βΒΫΒΡέœΜυpKa±δΜ·“‘ΦΑάμ¬έΦΤΥψΒΡΫαΙϊœύΈ«ΚœΘέ71Θ§72Θί. ÷ Ή”Μ·ΒΡB-Block HisΤπΒΫΥα¥ΏΜ·Ής”ΟΘ§ ‘Ύ÷ Ή”Μ·Κσ≥…ΈΣ«βΦϋΙ©ΧεΘ§ ”κθΘΑΖΦϋτ Μυ―θœύΜΞΉς”ΟΘ§ ‘ω«ΩΝΥτ ΜυΒΡ«ΉΒγ–‘. ’β“ΜœύΜΞΉς”ΟΒΟΒΫΝΥάμ¬έΦΤΥψΒΡ÷ß≥÷Θέ72ΘίΘ§ ≤ΔΫχ“Μ≤Ϋ‘ΎSsp DnaB M86ΡΎΚ§κΡΒΡΨßΧεΫαΙΙ÷–ΒΟΒΫΝΥ»Ζ»œΘέ73Θί. B-Block ThrΤπΒΫΦν¥ΏΜ·Ής”ΟΘ§ ”κθΘΑΖΦϋΒΡΑ±Μυ«βœύΜΞΉς”ΟΘ§ ΙθΘΑΖΦϋΤΪάκάμœκΦϋ≤Έ ΐΘ§ Ά®ΙΐΧα…ΐΒΉΈοΡήΝΩΫΒΒΆΝΥΜνΜ·Ρή. ’β“ΜΜζάμΒΟΒΫΝΥSsp DnaEΡΎΚ§κΡΨßΧεΫαΙΙΒΡ÷ß≥÷Θέ74ΘίΘ§ ΒψΆΜ±δ Β―ι±μΟςΗΟΑ±ΜυΥαΕ‘N-SθΘΜυ«®“Τ «±Ί“ΣΒΡΘέΆΦ7Θ®AΘ©ΘίΘέ75Θί.
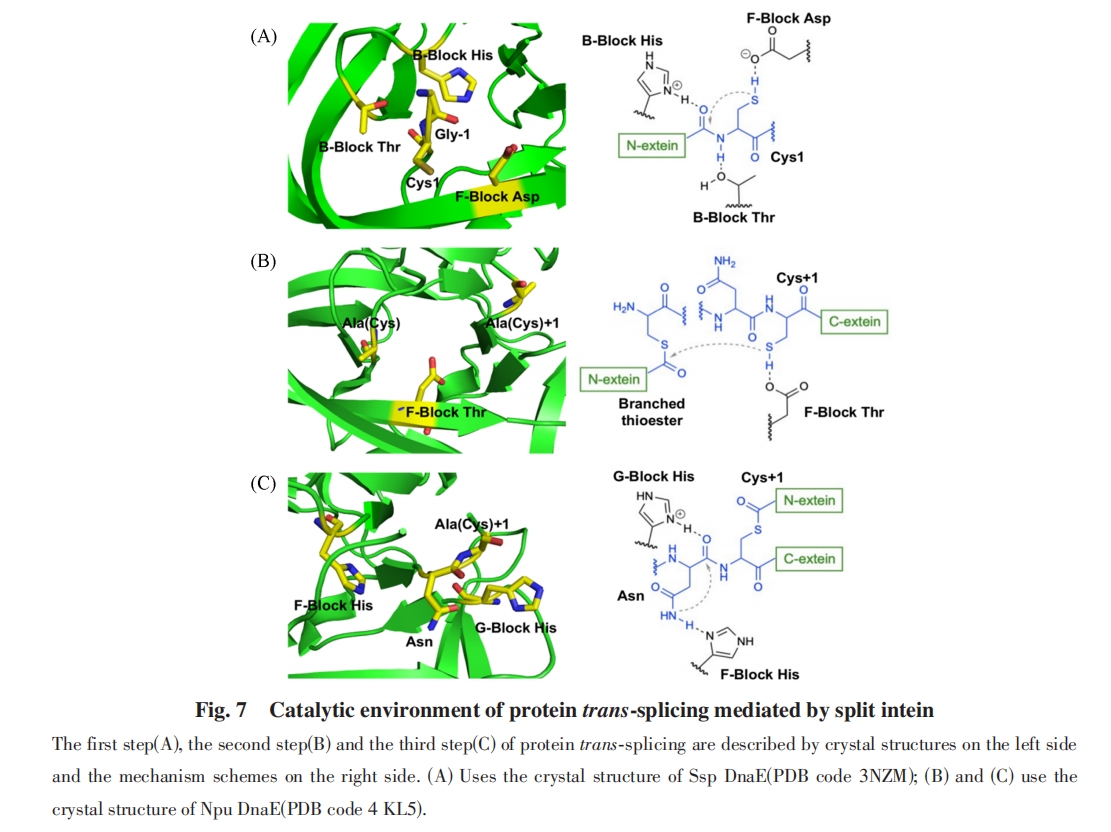
ΒΎΕΰ≤ΫΘ§ C-ΕΥΆβœ‘κΡΒΡCys≤ύΝ¥έœΜυΫχΙΞN-ΕΥΆβœ‘κΡ”κN-ΕΥΡΎΚ§κΡ÷°ΦδΒΡΝρθΞΦϋΘ§ Ά®ΙΐΝρθΞΫΜΜΜ ΙΝΫΕΈΆβœ‘κΡΆ®ΙΐΝρθΞΦϋœύΝ§Θ§ ¥”œΏ–‘ΝρθΞ÷–ΦδΧεΉΣΜ·ΈΣ÷ßΝ¥ΝρθΞ÷–ΦδΧε. QM/MMΦΤΥψΫαΙϊ±μΟςΘ§ F-Block Asp‘Ύ¥ΥΙΐ≥Χ÷–Ά§―υΤπΒΫΦν¥ΏΜ·ΒΡΉς”ΟΘ§ ¥ΌΫχΝΥC-ΕΥΆβœ‘κΡΒΡCysΒΡ»Ξ÷ Ή”Μ·. ΒψΆΜ±δ Β―ι±μΟςΘ§ ΫΪ’βΗωAspΆΜ±δΈΣGluΘ§ AsnΘ§ AlaΜρGlyΕΦΜαœ‘÷χ“÷÷Τ÷ßΝ¥÷–ΦδΧεΒΡ–Έ≥…ΘέΆΦ7Θ®BΘ©ΘίΘέ76Θί.
ΒΎ»ΐ≤ΫΘ§ C-ΕΥΡΎΚ§κΡΒΡC-ΕΥAsn≤ύΝ¥θΘΑΖΫχΙΞC-ΕΥΡΎΚ§κΡ”κC-ΕΥΆβœ‘κΡ÷°ΦδΝ§Ϋ”ΒΡθΘΑΖΦϋΘ§ –Έ≥…ΕΰθΘ―«ΑΖΈε‘ΣΜΖΘ§ ΡΎΚ§κΡ”κΆβœ‘κΡΆ―άκ. F-BlockΚΆG-Block÷–ΒΡ2ΗωHisΤπΒΫΝΥ¥ΏΜ·Ής”Ο. Τδ÷–Θ§ F-Block His¥ΌΫχΝΥC-ΕΥAsn≤ύΝ¥θΘΑΖΒΡ»Ξ÷ Ή”Μ·Θ§ ΤπΒΫΦν¥ΏΜ·Ής”Ο. ÷ Ή”Μ·ΚσΒΡG-Block His”κθΘΑΖΦϋτ Μυ―θœύΜΞΉς”ΟΘ§ ΤπΒΫΝΥ‘ω«Ωτ Μυ«ΉΒγ–‘ΒΡΥα¥ΏΜ·Ής”Ο. …œ ωœύΜΞΉς”Ο‘ΎSsp DnaBΒΡΨßΧεΫαΙΙ÷–ΒΟΒΫΝΥΙέ≤βΘέ77ΘίΘ§ “≤ΒΟΒΫΝΥΦΤΥψΫαΙϊΒΡ÷ß≥÷Θέ78Θί. ¥ΥΆβΘ§ MuirΒ»Θέ79ΘίΖΔœ÷Ε‘”ΎNpu DnaEΡΎΚ§κΡΘ§ ‘ΎΈόΖ®–Έ≥…÷ßΝ¥÷–ΦδΧεΒΡC1AΆΜ±δΧε÷–Θ§ AsnΜΖΜ·ΥΌ¬ ΜαΫΒΒΆ÷ΝΆΜ±δ«ΑΒΡ1/200Θ§ ΥΒΟς÷ßΝ¥÷–ΦδΧεΒΡ–Έ≥…¥ΌΫχΝΥAsnΒΡΜΖΜ·ΘέΆΦ7Θ®CΘ©Θί.
ΉνΚσΘ§ ‘ΎΡΎΚ§κΡ¥”ΧεœΒ÷–Ά―άκΚσΘ§ ΝΫΕΈΆβœ‘κΡΦδΒΡΝρθΞΦϋΆ®ΙΐS-NθΘΜυ«®“Τ–Έ≥…Χλ»ΜκΡΦϋΘ§ ΫχΕχΒΟΒΫΦτ«–≤ζΈο. ΗΟΙΐ≥Χ≤Μ–η“ΣΤδΥϋΗ®÷ζΘ§ ‘Ύ÷––‘ΧθΦΰœ¬ΡήΙΜΉ‘ΖΔΫχ––Θέ80Θί.
‘Ύ“Μ–©Ζ÷Ν―ΡΎΚ§κΡ÷–Θ§ …œ ωΜζάμ÷–ΒΡCys±ΜSerΚΆThrΧφ¥ζΘ§ Ζ¥”ΠΙΐ≥Χ÷––Έ≥…œύ”ΠΒΡ―θθΞ÷–ΦδΧε Θ®±μ1ΚΆ±μ2Θ©. ¥ΥΆβΘ§ ≤ΩΖ÷ΡΎΚ§κΡΒΎ“Μ≤ΫΦτ«–ΒΡ«ΉΚΥΑ±ΜυΥαΈΜ”ΎΡΎΚ§κΡΡΎ≤ΩΕχ≤Μ «NΕΥΘ§ MootzΒ»Θέ58Θί±®ΒάΝΥΒΎ“ΜΗωΉώ―≠’β÷÷ΜζάμΒΡΧλ»ΜΖ÷Ν―ΡΎΚ§κΡNrdHFΘ§ NrdHF≤Έ”κΒΎ“Μ≤ΫN-SθΘΜυ«®“ΤΒΡCysΈΜ”ΎIntC–ρΝ–ΒΡ÷–Φδ≤ΩΈΜ.
≥ΐΝΥΡΎΚ§κΡ–ρΝ–ΡΎ≤ΩΑ±ΜυΥαΒΡ¥ΏΜ·Ής”ΟΘ§ Ζ÷Ν―ΡΎΚ§κΡΜΙ±μœ÷≥ωΕ‘Άβœ‘κΡ–ρΝ–ΒΡΟτΗ––‘ΘΚΩΩΫϋΦτΫ”ΈΜΒψΒΡΆβœ‘Α±ΜυΥαΆΜ±δΕ‘Ζ¥”Π–ß¬ ΚΆ≤ζ¬ ”–Οςœ‘”ΑœλΘ§ ΑΒ ΨΆβœ‘–ρΝ–Ω…Ρή”κΙΊΦϋΑ±ΜυΥα¥φ‘ΎœύΜΞΉς”Ο. MuirΒ»Θέ81ΘίΫΪNpu*ΡΎΚ§κΡΘ®Ssp DnaE-NΚΆNpu DnaE-CΒΡΉιΚœΘ©C-ΕΥΆβœ‘κΡ¥”Χλ»ΜΒΡCFNΗΡΈΣSGVΘ§ ΖΔœ÷CΕΥΦτΫ”–߬ Οςœ‘œ¬ΫΒ. Ά®ΙΐΕ®œρΫχΜ·Θ§ ΒΟΒΫΝΥΩ…“‘Ηϋ”––ßΦτΫ”SGVΉςΈΣC-ΕΥΆβœ‘κΡΒΡmNpu*Θ§ Τδ÷–Αϋά®F-Block D124YΆΜ±δ. ΨßΧεΫαΙΙ―–ΨΩœ‘ ΨΘ§ CΕΥΆβœ‘κΡCFN÷–ΒΡPhe”κ¥ΌΫχAsnΜΖΜ·ΒΡF-Block His125¥φ‘ΎΠ–-Π–Ε―ΒΰΉς”ΟΘ§ ΙHis125¥Π”Ύ”–άϊ”Ύ¥ΏΜ·ΒΡ»ΓœρΘ§ ΕχD124YΆΜ±δ ΙTyr¥ζΧφΝΥPheΒΡΉς”ΟΘ§ ¥”ΕχΦθ«αΝΥΖ÷Ν―ΡΎΚ§κΡΕ‘C-ΕΥΆβœ‘κΡΒΡΟτΗ––‘. MootzΒ»Θέ82ΘίΖΔœ÷ΫΪSsp DnaB N-ΕΥΆβœ‘κΡΒΡGlyΗΡ±δΈΣL-AlaΜαΒΦ÷¬ΦτΫ”–߬ Οςœ‘œ¬ΫΒΘ§ ΕχΗΡ±δΈΣD-AlaΒΡ–ß¬ œ¬ΫΒΖυΕ»Ϋœ–Γ. Ά®ΙΐΙέ≤λΨßΧεΫαΙΙΘ§ »œΈΣN-Άβœ‘κΡΒΡΑ±ΜυΥα≤ύΝ¥Ω…Ρή”κB-Block His≤ύΝ¥≤ζ…ζΈΜΉη≥εΆΜΘ§ Η…»≈HisΒΡ¥ΏΜ·ΙΠΡήΘ§ ΕχΤδΥϋ≥·œρΒΡ≤ύΝ¥Θ®»γD-AlaΘ©≤ζ…ζΒΡΈΜΉη≥εΆΜΫœ–Γ. ’β–© Β―ιΫαΙϊΥΒΟςΆβœ‘κΡ”κΡΎΚ§κΡ÷°Φδ¥φ‘ΎΜΞΉςΘ§ Ω…Ρή «Άβœ‘–ρΝ–ΟτΗ––‘ΒΡ÷Ί“Σ‘≠“ρ.
3 Ζ÷Ν―ΡΎΚ§κΡΒΡΙΛ≥ΧΜ·ΗΡ‘λ”κ–‘Ρή”≈Μ·
Ζ÷Ν―ΡΎΚ§κΡ“―Ψ≠≥…ΈΣ“Μ÷÷ΙψΖΚ”Π”ΟΒΡΒΑΑΉ÷ Ν§Ϋ”ΙΛΨΏΘ§ ΒΪ‘ΎΤδ”Π”ΟΙΐ≥Χ÷–“ά»Μ¥φ‘Ύ“Μ–©Χτ’ΫΘ§ “ρ¥Υ―–ΨΩ’ΏœΘΆϊΡήΜώΒΟΨΏ”–Ηϋ”≈–‘ΡήΒΡΡΎΚ§κΡΘ§ Αϋά®Φτ«–Μν–‘ΚΆΈ»Ε®–‘ΗΏΓΔ Ε‘Άβœ‘κΡΦτΫ”ΈΜΒψΗΫΫϋΑ±ΜυΥα–ρΝ–ΒΡΡΆ ή–‘«ΩΘ§ ≤ΜΆ§Ζ÷Ν―ΡΎΚ§κΡΦδΒΡΖ¥”Π’ΐΫΜΘ§ Ζ÷Ν―ΡΎΚ§κΡ–ρΝ–ΕΧΘ§ C-ΕΥΜρN-ΕΥ≥ΛΕ»‘ΎΙΧœύΚœ≥…ΖΕΈßΡΎΒ». ΈΣΜώΒΟΨΏ”–…œ ωΧΊ–‘ΒΡΖ÷Ν―ΡΎΚ§κΡΘ§ ―–ΨΩ’ΏΟ«Ϋχ––ΝΥΙψΖΚΒΡΧΫΨΩ”κ≥Δ ‘.
3.1ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΦτ«–Μν–‘ΚΆΈ»Ε®–‘
2016ΡξΘ§ MuirΒ»Θέ83Θί Ι”Οάμ–‘…ηΦΤΖΫΖ®ΜώΒΟΝΥΨΏ”–«ΩΈ»Ε®–‘ΚΆΗΏΦτ«–Μν–‘ΒΡΖ÷Ν―ΡΎΚ§κΡ. Ά®ΙΐΖ÷ΈωDnaEΦ“ΉεΒΡNpuΘ®ΩλΘ©ΚΆSspΘ®¬ΐΘ©Ζ÷Ν―ΡΎΚ§κΡ÷°ΦδΦτΫ”–߬ ΒΡ≤ν“λΖΔœ÷Θ§ ¥σΕύ ΐΙΊΦϋ≤–Μυ‘ΎΩ’ΦδΫαΙΙ…œ”κΜν–‘ΈΜΒψ÷±Ϋ”ΝΎΫϋ. “‘’β–©ΙΊΦϋΑ±ΜυΥα≤–ΜυΉςΈΣΕ‘ΤκΈΜΒψΘ§ Ε‘73ΗωDnaEΦ“ΉεΒΡΡΎΚ§κΡΫχ––±»Ε‘Θ§ Ήν÷’ΒΟΒΫΝΥ“ΜΗω–¬–ΆΖ÷Ν―ΡΎΚ§κΡCfa. Cfaœ‘ Ψ≥ωΩλΥΌΒΡΒΑΑΉ÷ ΦτΫ”Μν–‘“‘ΦΑΗΏΒΡ»»Έ»Ε®–‘ΚΆ±δ–‘Έ»Ε®–‘. ‘Ύ30 Γφœ¬Θ§ CfaΒΡΦτ«–ΥΌ¬ ±»¥Υ«Α±®ΒάΒΡDnaEΦ“Ήε÷–ΉνΩλΒΡNpu DnaEΩλ2.5±ΕΘ§ Τδt1/2ΫωΈΣ20 s. Ά§ ±Θ§ CfaΕ‘±δ–‘ΧθΦΰΨΏ”–Κή«ΩΒΡΡΆ ή–‘Θ§ ‘Ύ3~4 mol/L GnΓΛHClΜρ8 mol/LΡρΥΊΒ»«Ω±δ–‘ΧεœΒ÷–ΕΦ±Θ≥÷ΝΦΚΟΒΡΦτΫ”Μν–‘Θ§ ΦτΫ”Ζ¥”ΠΤΫΚβ≥Θ ΐK>1ΓΝ10-2 s-1.
3.2ΓΓΖ÷Ν―ΡΎΚ§κΡΕ‘ΦτΫ”ΈΜΒψΒΡΦφ»ί–‘
2011ΡξΘ§ LiuΒ»Θέ84ΘίΆ®ΙΐΕ®œρΫχΜ·ΦΦ θΘ§ ΒΟΒΫΝΥΨΏ”–ΗΏΆβœ‘κΡΦτΫ”ΈΜΒψΦφ»ί–‘ΒΡΡΎΚ§κΡ. ΫΪSsp DnaBΡΎΚ§κΡΜυ“ρ≤ε»κΒΫΩ®Ρ«ΟΙΥΊΩΙ–‘ΒΑΑΉ÷ Θ®KanRΘ©ΒΡΜυ“ρ÷–Θ§ ΙΒΟΚ§ΡΎΚ§κΡΒΡKanRΒΑΑΉ«ΑΧεΟΜ”–Ω®Ρ«ΟΙΥΊΩΙ–‘Θ§ ÷Μ”–ΖΔ…ζΝΥΒΑΑΉΦτΫ”Ζ¥”ΠΒΟΒΫΆξ’ϊKanRΒΑΑΉΘ§ Ω®Ρ«ΟΙΥΊΩΙ–‘Μ÷Η¥ΒΡΨζ÷ξ≤≈Ρή‘ΎΚ§Ω®Ρ«ΟΙΥΊΒΡ≈ύ―χΜυ÷–¥φΜν. Ε‘¥φΜνΨζ÷ξΒΡ÷ ΝΘΫχ––≤β–ρΘ§ Φ¥Ω…ΜώΒΟΡήΙΜ‘ΎΗΟ≤ε»κΈΜΒψΖΔ…ζΦτΫ”Ζ¥”ΠΒΡΡΎΚ§κΡΜυ“ρ–ρΝ–Θ®ΆΦ8Θ©. Ψ≠ΙΐΕύΗω…Η―Γ―≠ΜΖΘ§ ΫχΜ·ΒΟΒΫΝΥΕύΗω–¬–ΆΡΎΚ§κΡΘ§ ΡήΙΜ‘ΎΕύΗω≤ΜΆ§ΒΡΆβœ‘κΡΦτΫ”ΈΜΒψΚΆ≤ΜΆ§ΒΡΒΑΑΉ÷ ÷–ΕΦœ‘ Ψ≥ωΗΏΦτΫ”Μν–‘. Τδ÷–Θ§ M86ΆΜ±δΧεΡΎΚ§κΡΨΏ”–ΉνΚΟΒΡΦτΫ”Ζ¥”Π–‘Ρή. ΫΪM86Ά®Ιΐ»ΥΙΛΖ÷Ν―ΈΣNΕΥ”…11ΗωΑ±ΜυΥαΉι≥…ΒΡΖ÷Ν―ΡΎΚ§κΡ. ”κΆΜ±δ«ΑΒΡΖ÷Ν―DnaBΡΎΚ§κΡœύ±»Θ§ M86¥ΏΜ·ΒΑΑΉ÷ Ζ¥ ΫΦτΫ”Ζ¥”ΠΒΡΥΌ¬ ‘ωΦ”ΝΥ60±ΕΘ§ Ζ¥”Π≥Θ ΐkM86=2.5ΓΝ10-3 s-1Θ§ ≤Δ«“Ζ÷Ν―ΡΎΚ§κΡΤ§ΕΈΒΡKd÷ΒΧαΗΏΝΥ“ΜΗω ΐΝΩΦΕ.
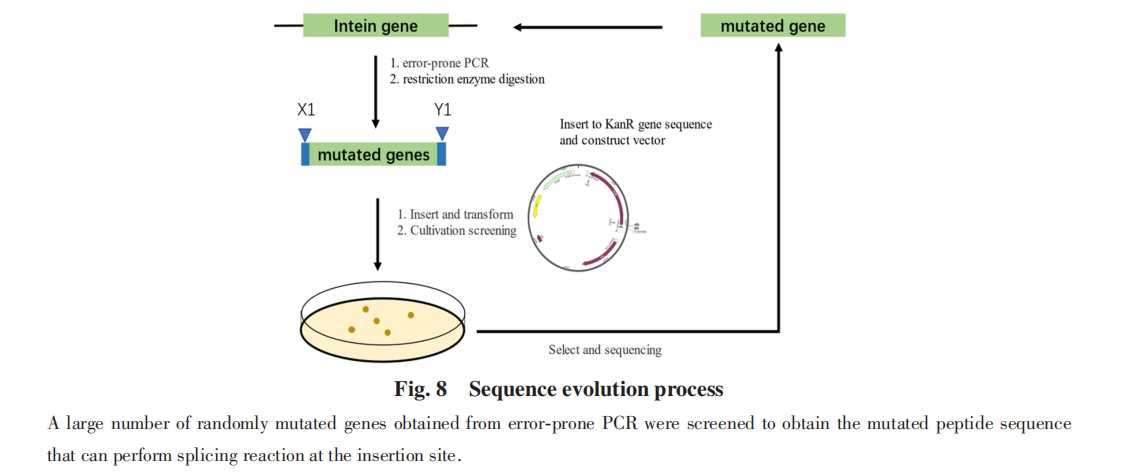
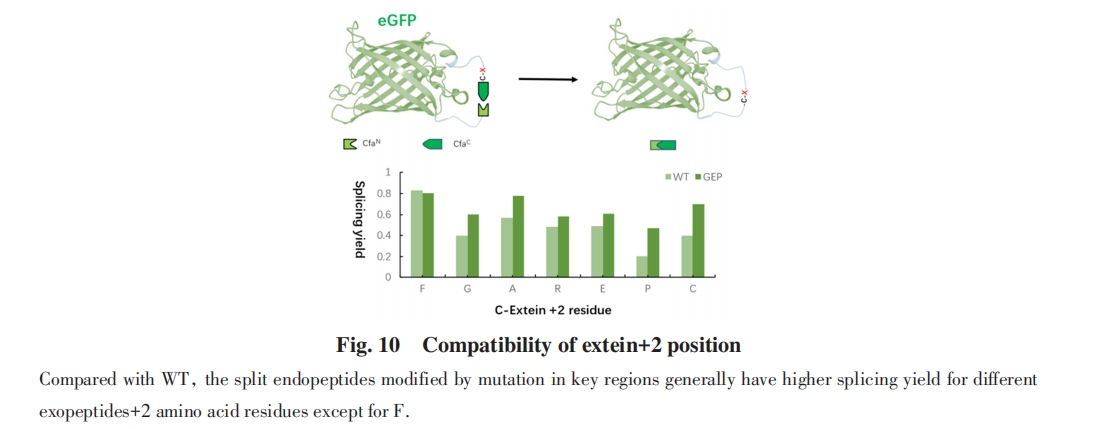
2017ΡξΘ§ MuirΒ»Θέ85Θί±®ΒάΝΥΆβœ‘κΡ–ρΝ–ΡΆ ή–‘Οςœ‘ΧαΗΏΒΡΉ‘»ΜΖ÷Ν―ΡΎΚ§κΡ. His125 «ΒΑΑΉ÷ ΦτΫ”ΉνΚσ“Μ≤Ϋ÷–Asn137ΜΖΜ·ΒΡΙΊΦϋ¥ΏΜ·≤–ΜυΘ§ Ά®Ιΐ…ηΦΤHis125Υυ‘ΎΒΡΜΖΘ®≤–Μυ122~124Θ©Θ§ Ω…“‘”––ßΒΊΗΡ±δHis125ΙΙœσΕ·ΝΠ―ßΘ§ ¥”Εχœ‘÷χ‘ωΦ”Φτ«–Ζ¥”ΠΕ‘+2”κ+3ΈΜ÷ΟΒΡΆβœ‘κΡ–ρΝ–ΡΆ ή–‘Θ§ ≤ΔΧαΗΏΝΥΦτ«–ΥΌ¬ . Μυ”ΎΕ‘ΒΑΑΉ÷ ΦτΫ”ΙΊΦϋΒΡ¥ΏΜ·ΈΜΒψΒΡάμΫβΘ§ MuirΒ»Ε‘Ζ÷Ν―ΡΎΚ§κΡHis125Υυ‘ΎΒΡΜΖΫχ––ΝΥ±ΞΚΆΆΜ±δΘ§ »ΜΚσ Ι”ΟΜυ”ΎœΗΑϊΒΡ―Γ‘ώœΒΆ≥ά¥ΦχΕ®ΡήΙΜ‘Ύ≤ΜΆ§Άβœ‘κΡΦτΫ”ΈΜΒψœ¬¥ΏΜ·”––ßΦτΫ”ΒΡΆΜ±δΧεΘ§ ΖΔœ÷¥σΕύ ΐΨζ¬δ‘Ύ≤–Μυ122~124¥ΠΚ§”–GXPΜυ–ρΘ®GRPΘ§ GEPΜρGPPΘ§ ¥ζΧφΧλ»ΜERD–ρΘ©Θ§ Τδ÷–GEP–ρΝ–ΨΏ”–ΉνΗΏΒΡΦτΫ”Μν–‘Θ®ΆΦ9Θ©.
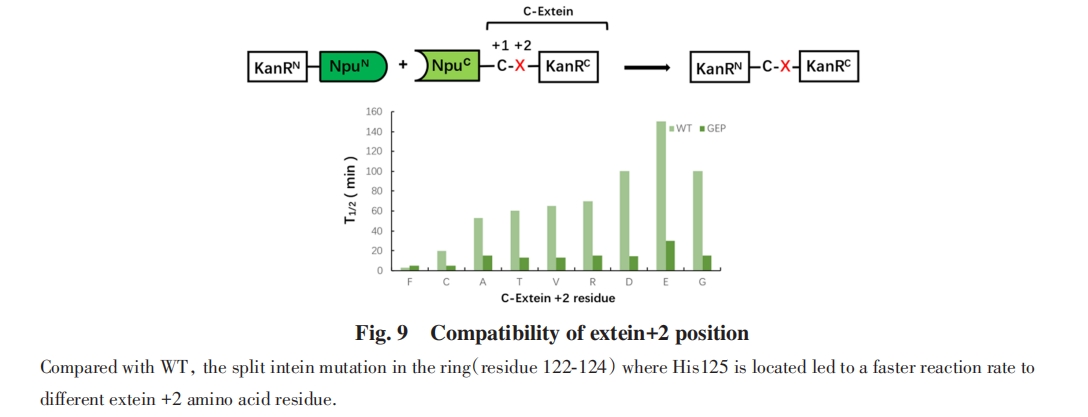
―–ΨΩ±μΟςΘ§ Npu DnaEΡΎΚ§κΡΒΡΆβœ‘κΡ–ρΝ–ΤΪΚΟ÷ς“ΣΨ÷œό”ΎCΕΥΆβœ‘κΡ+1ΈΜΒΡ¥ΏΜ·ΑκκΉΑ±ΥαΚΆ+2ΈΜΒΡ¥σΈΜΉη ηΥ°≤–ΜυΘ§ ΫΪΧλ»Μ+2ΈΜPheΆβœ‘κΡ–ρΝ–≤–ΜυΆΜ±δΈΣΧεΜΐΫœ–ΓΒΡ≤–ΜυΘ®άΐ»γAlaΘ©ΜαΒΦ÷¬ΦτΫ”¬ œ‘÷χΫΒΒΆΘέ79Θί. MuirΒ»Θέ85Θί“‘GFPΒΑΑΉΉςΈΣΡΘ–ΆΘ§ ±μ’ςΝΥΕ‘”Ύ≤ΜΆ§Άβœ‘κΡΈΜΒψΒΡΦτΫ”Μν–‘. Ψ≠ΙΐGEP–ρΝ–ΗΡ‘λΒΡNpu DnaEΡΎΚ§κΡ”κΈ¥ΗΡ‘λΒΡNpu DnaEΡΎΚ§κΡœύ±»Θ§ Ε‘≤ΜΆ§ΒΡ+2ΚΆ+3ΈΜΆβœ‘κΡ–ρΝ–ΒΡΦτΫ”≤ζ¬ Ψυ”–œ‘÷χΧαΗΏΘ®ΆΦ10Θ©.
3.3ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡΖ¥”Π’ΐΫΜ–‘
Ζ÷Ν―ΡΎΚ§κΡΒΡNΕΥΚΆCΕΥΤ§ΕΈ÷°ΦδΨΏ”–«ΩΝ“ΒΡΒγΚ…-ΒγΚ…œύΜΞΉς”Ο. 2019ΡξΘ§ IwaïΒ»Θέ86ΘίΜυ”ΎΕ‘Ζ÷Ν―ΡΎΚ§κΡ2ΗωΤ§ΕΈΫαΚœΙΐ≥ΧΓΑ≤ΕΜώΚΆ’έΒΰΜζ÷ΤΓ±÷–ΒΡΨ≤ΒγœύΜΞΉς”ΟΘ§ ΫΪ2ΗωΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΫαΚœΙΊΦϋΈΜ÷ΟΒΡΑ±ΜυΥαΒγΚ…Άχ¬γΫχ––ΗΡ‘λΘ§ Βœ÷ΝΥ2÷÷DnaBΦ“ΉεΖ÷Ν―ΡΎΚ§κΡΫαΚœΒΡ’ΐΫΜ–‘. ’β“Μ≤Ώ¬‘“≤ ”Ο”Ύ¥”ΤδΥϋΥ≥ ΫΦτΫ”ΡΎΚ§κΡ÷–≤ζ…ζ–¬ΒΡΖ÷Ν―ΡΎΚ§κΡ.
2020ΡξΘ§ WangΒ»Θέ87Θί…Η―ΓΦλ≤βΝΥΧλ»Μ¥φ‘ΎΒΡΖ÷Ν―ΡΎΚ§κΡΒΡΦτΫ”Ζ¥”ΠΘ§ Ϋ®ΝΔΝΥΑϋΚ§15÷÷Ζ÷Ν―ΡΎΚ§κΡΒΡ’ΐΫΜΖ÷Ν―ΡΎΚ§κΡΩβ. ―–ΨΩΫαΙϊ±μΟςΘ§ ’ΐΫΜΖ÷Ν―ΡΎΚ§κΡΩ…“‘”κΕύΗωΖ÷Ν―ΉΣ¬Φ“ρΉ”ΫαΚœΘ§ “‘‘ΎΜν…ζΈοΧε÷– Βœ÷Η¥‘”ΒΡ¬ΏΦ≠±μ¥οΙΐ≥ΧΘ§ ΜΙΩ…“‘”Ο”ΎΕύΤ§ΕΈΒΑΑΉΒΡΧεΆβΝ§Ϋ”. Ής’Ώ“‘SasGΒΑΑΉ÷ΊΗ¥Τ§ΕΈΫχ––’Ι ΨΘ§ Ι”Ο’ΐΫΜΖ÷Ν―ΡΎΚ§κΡΉιΚœΘ§ “ΜΙχΆξ≥…ΝΥ6ΗωΤ§ΕΈΒΡΤ¥Ϋ”Θ®ΆΦ11Θ©Θ§ ÷ΛΟςΝΥ’ΐΫΜΖ÷Ν―ΡΎΚ§κΡά©’ΙΩβΒΡΕύΙΠΡή–‘ΚΆΨό¥σ«±ΝΠ.

ΨΓΙήΖ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡPTS‘ΎΒΑΑΉ÷ ΑκΚœ≥……œ»ΓΒΟΝΥ“ΜΕ®ΒΡ≥…ΙΠΘ§ ΒΪ“≤¥φ‘Ύ“Μ–©Ψ÷œόΘ§ »γ–ό ΈΜυΆ≈ΒΡ―Γ‘ώ–‘“ΐ»κΈΜΒψΆ®≥Θ÷ΜΡήΈΜ”ΎΡΩ±ξΒΑΑΉΒΡNΕΥΜρCΕΥ. ΥφΉ≈‘Ϋά¥‘ΫΕύΒΡΖ÷Ν―ΡΎΚ§κΡ±ΜΦχΕ®Μρ…ηΦΤ≥ωά¥Θ§ Ζ÷Ν―ΡΎΚ§κΡΙΛΨΏœδ÷πΫΞΖαΗΜΘ§ ‘ΎΆ§“ΜΡΩ±ξΒΑΑΉΒΡΑκΚœ≥…÷–Ά§ ±άϊ”Ο2÷÷œύΜΞ’ΐΫΜΒΡΖ÷Ν―ΡΎΚ§κΡΫχ––Εύ¥ΈPTSΒΡ≤Ώ¬‘≥…ΈΣΩ…Ρή.
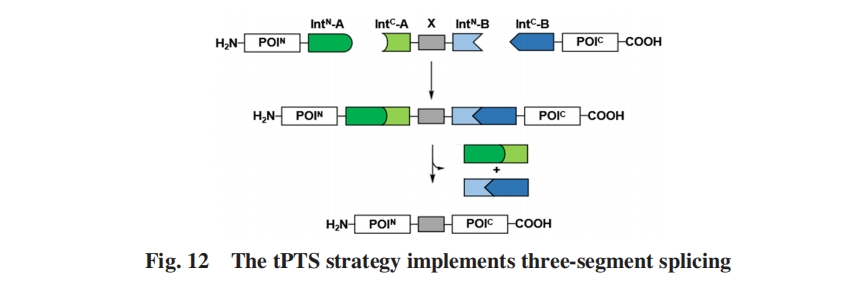
MuirΒ»Θέ88Θί Ή¥ΈΩΣΖΔΝΥΆ®Ιΐ2÷÷’ΐΫΜΖ÷Ν―ΡΎΚ§κΡ Βœ÷“ΜΙχ»ΐΤ§ΕΈΖ¥ ΫΦτΫ”ΒΡ≤Ώ¬‘Θ§ ΗΟΖΫΖ®±Μ≥ΤΈΣ¥°ΝΣΒΑΑΉΖ¥ ΫΦτΫ”Θ®tPTSΘ©. ΡΩ±ξΒΑΑΉΖ÷ΈΣ3ΗωΤ§ΕΈΘ§ Φ¥POINΚΆPOIC“‘ΦΑ“ΜΗωΆ®Ιΐ±μ¥οΜρΜ·―ßΚœ≥…Μώ»ΓΒΡ¥χ”–Υυ–η–ό ΈΜυΆ≈ΒΡΫœΕΧ÷––ΡΕύκΡΤ§ΕΈXΘ®ΆΦ12Θ©. Υυ Ι”ΟΒΡ2Ηω’ΐΫΜΖ÷Ν―ΡΎΚ§κΡΘ®AΚΆBΘ©”κPOINΚΆ POIC»ΎΚœ±μ¥οΘ®POIN+IntN-AΦΑIntC-B+POICΘ©Θ§ ΕχX”κAΚΆBΒΡΝμΆβ2ΗωΤ§ΕΈ»ΎΚœΘ®IntC-A+X+IntN-BΘ©. ’β÷÷–¬”±ΒΡœΒΆ≥Ω…“‘ Βœ÷3ΗωΕύκΡΤ§ΕΈΒΡ“ΜΙχΜ·ΉιΉΑΘ§ Κœ≥…ΝΥ304Ηω≤–Μυ¥σ–ΓΒΡc-CrkIIΒΑΑΉΘ§ Τδ”…“ΜΗωSH2ΫαΙΙ”ρΚΆ2ΗωSH3ΫαΙΙ”ρΉι≥…Θ§ ΟΩΗωΫαΙΙ”ρΖ÷±π”κœύ”ΠΖ÷Ν―ΡΎΚ§κΡΙΙ≥…tPTSΒΡ«ΑΧεΤ§ΕΈ. Ζ¥”Π‘Ύ22 Γφœ¬Ϋχ––ΝΥ48 hΘ§ “‘‘Φ48%ΒΡ ’¬ Άξ≥…ΝΥ»ΐΤ§ΕΈΦτΫ”. ΗΟΖΫΖ®ΈΣΫœ¥σΒΡΕύΫαΙΙ”ρΒΑΑΉΒΡΑκΚœ≥…ΦΑΒΑΑΉ–ρΝ–÷–―κΧΊΕ®ΈΜΒψ“ΐ»κΜ·―ß–ό ΈΧαΙ©ΝΥ“Μ÷÷ΖΫ±ψΒΡΖΫΖ®.
3.4ΓΓΖ÷Ν―ΡΎΚ§κΡΒΡ Ι”Ο±ψΫί–‘
Χλ»ΜΕœΝ―ΒΡgp41-1ΡΎΚ§κΡ «ΡΩ«Α±®ΒάΒΡΉν–ΓΒΡΖ÷Ν―ΡΎΚ§κΡ÷°“ΜΘ§ ΨΏ”–Ζ«≥ΘΗΏΒΡΖ¥ ΫΦτΫ”Μν–‘Θέ56ΘίΘ§ gp41-1ΡΎΚ§κΡ”…88Ηω≤–ΜυΒΡIntNΚΆ37Ηω≤–ΜυΒΡIntCΉι≥…. ΈΣΝΥ ΙΡΎΚ§κΡΕ‘Άβœ‘κΡ–‘÷ ΒΡΗ…»≈Ηϋ–ΓΘ§ »ΥΟ«Ά®≥ΘœΘΆϊ ΙΡΎΚ§κΡΨΓΩ…Ρή–Γ. IwaïΒ»Θέ86Θί≥Δ ‘¥”gp41-1ΡΎΚ§κΡΒΡΉ‘»ΜΖ÷Ν―ΈΜΒψ¥ΠΫΊΕΧ2Ηω≤–ΜυΘ§ ¥”ΕχΜώΒΟΗϋ–ΓΒΡΖ÷Ν―ΡΎΚ§κΡ. »ΜΕχΘ§ ’β÷÷»± ß ΙΡΎΚ§κΡΒΡΒΑΑΉ÷ ΦτΫ”–߬ ΫΒΒΆΝΥ‘Φ40%Θ§ Ής’ΏΚσ–χ ‘ΆΦ”≈Μ·ΡΎΚ§κΡΒΡΑ±ΜυΥα–ρΝ–“‘Μ÷Η¥ΫΊΕΧΒΡgp41-1ΡΎΚ§κΡΒΡΦτΫ”Μν–‘ΒΫ‘≠”–Υ°ΤΫΘ§ ΒΪΟΜ”–≥…ΙΠ. ’β±μΟςgp41-1ΡΎΚ§κΡΒΡ¥σ–ΓΩ…Ρή“―¥οΒΫ Βœ÷ΙΠΡήΒΡΉν–Γ÷Β.
NeqΘ§ NpuΚΆSsp DnaEΒ»¥σ≤ΩΖ÷Χλ»ΜΖ÷Ν―ΡΎΚ§κΡΕΦ‘Ύ≥ΘΙφΒΡΡΎΚ§κΡΖ÷Ν―ΈΜΒψS0¥ΠΕœΝ―Θ§ ΗΟΈΜΒψ“≤≥δΒ±Ιι≥≤ΚΥΥαΡΎ«–ΟΗ≤ε»κΒΡ±ξΉΦΈΜ÷Ο. ΗΟΈΜΒψΩΩΫϋΡΎΚ§κΡCΕΥΘ§ ΕœΝ―≤ζ…ζ‘Φ100ΗωΑ±ΜυΥα≤–ΜυΒΡIntNΚΆ35ΗωΑ±ΜυΥα≤–ΜυΒΡIntC. IntCΫœΕΧΘ§ ±ψ”Ύ”κCΕΥΆβœ‘κΡΤ§ΕΈΙ≤Ά§ΙΧœύΚœ≥…Θ§ “ρ¥Υ’β―υΒΡΖ÷Ν―ΡΎΚ§κΡΈΣΒΑΑΉ÷ CΕΥΑκΚœ≥…“‘ΦΑ±ξΦ«ΧαΙ©ΝΥΖΫ±ψ. »ΜΕχΘ§ ”…”ΎΒΑΑΉ÷ ΙΧœύΚœ≥…ΡήΝΠΒΡœό÷ΤΘ§ Ά®ΙΐΙΧœύΚœ≥…”κN-ΡΎΚ§κΡœύΝ§ΒΡΒΑΑΉ «œύΕ‘άßΡ―ΒΡΘ§ ’β“≤œό÷ΤΝΥΖ÷Ν―ΡΎΚ§κΡ‘ΎNΕΥΑκΚœ≥…ΒΑΑΉ÷ Μρ’ΏΒΑΑΉ÷ NΕΥ–ό ΈΒ»≥ΓΨΑΒΡ”Π”Ο. “ρ¥ΥΘ§ ―–ΨΩ’ΏΟ«œΘΆϊ―Α’“ΜρΩΣΖΔΨΏ”–ΗϋΕΧNΕΥΤ§ΕΈΒΡΖ÷Ν―ΡΎΚ§κΡ.
2004ΡξΘ§ LiuΒ»Θέ63ΘίΫΪSsp DnaBΟ‘ΡψΡΎΚ§κΡ¥”Ζ«≥ΘΙφΒΡΖ÷Ν―ΈΜΒψ«–ΗνΘ§ ±®ΒάΝΥ3Ηω–¬ΒΡΖ÷Ν―ΈΜΒψS1Θ§ S8ΚΆS11Θ§ ¥”’β3÷÷ΈΜΒψ«–ΕœΘ§ ΕΦ≤ζ…ζΝΥΡήΙΜΫχ––ΒΑΑΉ÷ Ζ¥ ΫΦτΫ”ΒΡΝΫΕΈ ΫΖ÷Ν―ΡΎΚ§κΡΘ®ΆΦ13Θ©. Τδ÷–S1Ζ÷Ν―ΡΎΚ§κΡΒΡNΕΥΫω”–11ΗωΑ±ΜυΥα. ”…”Ύ≤ΜΆ§ΒΡΡΎΚ§κΡΨΏ”–œύΥΤΒΡΫαΙΙΘ§ ’β–©–¬ΒΡΖ÷Ν―ΈΜΒψΩ…“‘ΆΤΙψΒΫΤδΥϋΡΎΚ§κΡ. LiuΒ»Θέ63ΘίΜΙ Ή¥Έ÷ΛΟςΝΥΖ÷Ν―ΈΣ3ΕΈΒΡΖ÷Ν―ΒΡΡΎΚ§κΡ“ά»ΜΩ…“‘Ήι≥…Η¥ΚœΈο≤ΔΜ÷Η¥ΒΑΑΉ÷ ΦτΫ”Μν–‘.

2014ΡξΘ§ MootzΒ»Θέ59Θί¥”ΚξΜυ“ρΉι ΐΨί÷–ΦχΕ®≤Δ±μ’ςΝΥ–¬ΒΡΖ÷Ν―ΡΎΚ§κΡAceL-TerL. ’β « Ή¥ΈΖΔœ÷¥φ‘ΎΧλ»ΜΖ«Βδ–ΆΖ÷Ν―ΒΡΡΎΚ§κΡ. ÷Μ”–25ΗωΑ±ΜυΥαΒΡNΕΥΤ§ΕΈ «ΤυΫώΈΣ÷ΙΉνΕΧΒΡΧλ»ΜIntNΤ§ΕΈΘ§ “Ή”ΎΆ®ΙΐΙΧœύΚœ≥…Μώ»Γ. ‘Ύ8 ΓφΒΡΒΆΈ¬œ¬Θ§ ΗΟΡΎΚ§κΡΨΏ”–ΉνΗΏ¥ο90%ΒΡΒΑΑΉ÷ Ζ¥ ΫΦτΫ”–߬ . Ής’ΏΆ®ΙΐΕ®œρΒΑΑΉ÷ ΫχΜ·―Γ‘ώΝΥΫχ“Μ≤ΫΗΡΫχΒΡΆΜ±δΧε. ΙΛ≥ΧΜ·ΒΡΡΎΚ§κΡΆΜ±δΧε‘Ύ37 Γφœ¬ΨΏ”–ΗϋΗΏΒΡΦτΫ”–߬ Θ§ Ζ¥”ΠΥΌ¬ ≥Θ ΐk>1.8ΓΝ10-3 s-1Θ§ ≤Δ‘ΎΜ·―ß±ξΦ«Εύ÷÷ΒΑΑΉ÷ ”Π”Ο÷–’Ιœ÷≥ωΧλ»Μ–ρΝ–ΉνΗΏ50±ΕΒΡ–ß¬ .
4 Ζ÷Ν―ΡΎΚ§κΡ‘ΎΒΑΑΉ÷ άκΧε”κ‘ΎΧεΚœ≥…÷–ΒΡ”Π”Ο
4.1ΓΓάκΧεΒΑΑΉ÷ ΑκΚœ≥…
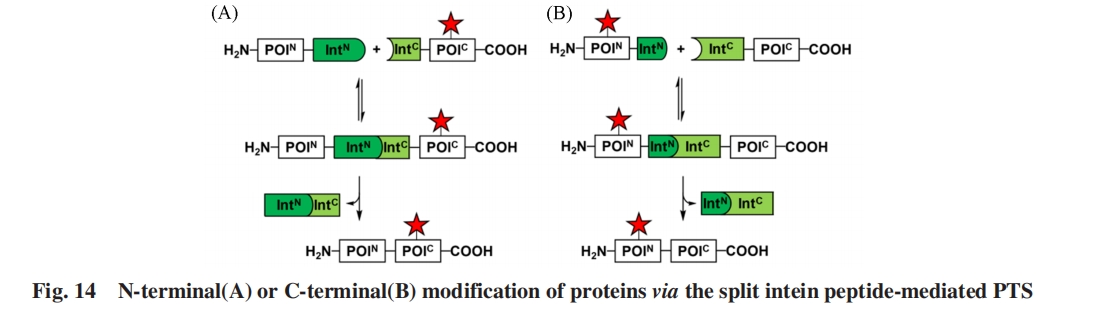
Ζ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡPTSΈΣΒΑΑΉ÷ ΑκΚœ≥…ΧαΙ©ΝΥ“Μ÷÷–¬ΆΨΨΕ. ≤ΜΆ§”ΎNCLΘέ15ΘίΚΆ±μ¥οΒΑΑΉΝ§Ϋ” Θ®EPLΘ©Θέ89~92ΘίΒ»±ξΉΦΒΑΑΉ÷ Μ·―ßΝ§Ϋ”Ζ¥”Π–η“ΣΫœΗΏ≈®Ε»Θ®“ρΤδ“άάΒ”ΎΕύκΡΤ§ΕΈΒΡΥφΜζ≈ωΉ≤Θ©Θ§ PTSΡήΙΜ‘ΎΠΧmol/L≈®Ε»ΧθΦΰœ¬Ζ¥”ΠΘ§ œ‘ Ψ≥ωΫœΒΆ≈®Ε»“άάΒ–‘Θ®“ρΤδ”…2ΗωΤ§ΕΈ÷°ΦδΫτΟήΒΡΒΑΑΉ÷ -ΒΑΑΉ÷ œύΜΞΉς”Ο¥Ό≥…Θ©Θέ69ΘίΘ§ ’β ΙΒΟΖ÷Ν―ΡΎΚ§κΡ≥…ΈΣΒΑΑΉ÷ ΑκΚœ≥…ΒΡ”–Έϋ“ΐΝΠΒΡΙΛΨΏ.
Ά®ΙΐPTSΆξ≥…ΑκΚœ≥…ΒΡΡΩ±ξΒΑΑΉΘ®POIΘ©ΫΪ±Μ≤πΖ÷ΈΣNΕΥΘ®POINΘ©”κCΕΥΘ®POICΘ©2Ηω«ΑΧεΤ§ΕΈΘ§ ΤδΉςΈΣΆβœ‘κΡΖ÷±π”κœύ”ΠΒΡΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈœύΝ§Ϋ”Θ§ ”…¥ΥΉι≥…ΒΡ»ΎΚœΧεΤ§ΕΈΖ÷±πΕΦΩ…“‘‘ΎPTSΖ¥”Π÷°«ΑΒΞΕά÷Τ±Η≤ΔΫΪΥυ–ηΒΡ–ό Έ“ΐ»κœύ”ΠΒΡΤ§ΕΈ÷–. ”…”Ύ–μΕύΨ≠Ιΐ”≈Μ·ΒΡΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΫœ–ΓΘ§ Ι Κ§”–Ϋœ–ΓΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΒΡΡΩ±ξΒΑΑΉ«ΑΧεΤ§ΕΈΘ®POIN⁃IntNΜρ IntC⁃POICΘ©Ω…Ά®ΙΐΙΧœύΕύκΡΚœ≥…ΜώΒΟΘ§ ¥”ΕχΩ…“‘»ί“ΉΒΊΫΪ–ό ΈΜυΆ≈―Γ‘ώ–‘“ΐ»κΡΩ±ξΒΑΑΉΒΡN-ΕΥΜρC-ΕΥ.
MootzΒ»Θέ11ΘίΫΪSsp DnaBΖ÷Ν―ΡΎΚ§κΡ”Ο”ΎΒΑΑΉ÷ ΑκΚœ≥…“‘ Βœ÷NΕΥ–ό Έ. ΗΟIntNΤ§ΕΈΫωΚ§11ΗωΑ±ΜυΥαΘ§ ±ψ”ΎΆ®ΙΐΙΧœύΕύκΡΚœ≥…Ά§ ±÷Τ±ΗIntN”κΚ§–ό ΈΜυΆ≈ΒΡΡΩ±ξΒΑΑΉΤ§ΕΈΘέΆΦ14Θ®AΘ©Θί. ΈΣ÷ΛΟςPTS‘ΎN-ΕΥ–ό ΈΒΑΑΉ÷ ΑκΚœ≥…÷–ΒΡ–ßΙϊΘ§ Ε‘2÷÷ΒΑΑΉ÷ ΒΉΈοΫχ––ΝΥN-ΕΥ”ΪΙβΥΊ±ξΦ«. Κœ≥…κΡ5-Fl-SEFSG-IntN”κ»ΎΚœ±μ¥οΒΡΒΑΑΉΤ§ΕΈIntC-Trx-His6“‘Β»ΡΠΕϊ≈®Ε»Θ®62 ΠΧmol/LΘ©‘Ύ25 Γφœ¬Ζ¥”Π22 hΘ§ Ψ≠¥ΩΜ·ΚσΒΡΑκΚœ≥…≤ζΈοFl-Trx-His6Ήή ’¬ ‘ΦΈΣ30%. ΒΎΕΰΗωάΐΉ” «Ζ÷Ή”ΝΩ30700ΒΡΠ¬-ΡΎθΘΑΖΟΗΘ®Π¬LacΘ©. »ΎΚœ±μ¥οΒΑΑΉΤ§ΕΈIntC-Π¬Lac-His6”κ5-Fl-SEFSG-IntN‘Ύ25 ΓφΘ§ 12 ΠΧmol/L≈®Ε»œ¬Ι≤Ζθ”ΐ42 hΘ§ “‘‘Φ30%ΒΡ ’¬ ΒΟΒΫΦτΫ”≤ζΈο Fl-Π¬Lac-His6.
LiuΒ»Θέ93Θί±®ΒάΝΥ“Μ÷÷–¬–ΆΙΛ≥ΧΜ·S11 Ssp GyrBΖ÷Ν―ΡΎΚ§κΡ. ΤδΖ÷Ν―ΈΜΒψ‘ΎC-ΕΥΗΫΫϋΘ§ ≤ζ…ζ≥ΛΕ»‘Φ150ΗωΑ±ΜυΥα≤–ΜυΒΡIntNΤ§ΕΈΚΆ≥ΛΕ»ΫωΈΣ6ΗωΑ±ΜυΥα≤–ΜυΒΡIntCΤ§ΕΈΘ§ Ω…”Ο”ΎΫΪΚœ≥…κΡΝ§Ϋ”ΒΫ±μ¥οΒΑΑΉΒΡCΕΥΘέΆΦ14Θ®BΘ©ΘίΘέ94Θί. ’β–©–¬”±ΒΡΑϋΚ§Ζ«≥ΘΕΧΒΡNΕΥΜρCΕΥΤ§ΕΈΒΡΖ÷Ν―ΡΎΚ§κΡœ‘÷χά©’ΙΝΥΤδ‘ΎΒΑΑΉ÷ ΑκΚœ≥…÷–ΒΡ”Π”ΟΘ§ ΙΒΟΫΪ–ό ΈΜυΆ≈ΧμΦ”ΒΫΡΩ±ξΒΑΑΉΒΡNΕΥΜρCΕΥΗϋΦ”ΖΫ±ψ.
άϊ”ΟΖ÷Ν―ΡΎΚ§κΡΜΙΩ…“‘ Βœ÷ΒΑΑΉ÷ ΒΡΈΜΒψ―Γ‘ώ–‘–ό Έ. ΈΣΝΥΩΥΖΰΕ‘Κ§”–ΕύΗωΑκκΉΑ±ΥαΒΡΡΩ±ξΒΑΑΉΡ―“‘ Βœ÷―Γ‘ώ–‘–ό ΈΒΡœό÷ΤΘ§ Φ¥‘ΎΤδ÷–“ΜΗωΧΊΕ®ΑκκΉΑ±ΥαΈΜΒψΒΡ≤ύΝ¥έœΜυ“ΐ»κ–ό ΈΕχ≤Μ”ΑœλΤδ”ύΑκκΉΑ±Υα≤–ΜυΘ§ MootzΒ»Θέ13ΘίΫΪΑκκΉΑ±Υα–ό Έ”κPTSΫαΚœ Ι”Ο. ΨΏΧεά¥ΥΒΘ§ »ΥΙΛΖ÷Ν―ΒΡSsp DnaBΚΆMxe GyrAΒΡIntCΤ§ΕΈΒΡΆβœ‘κΡ+1ΈΜΒψΖ÷±π”…ΥΩΑ±ΥαΜρΥ’Α±ΥαΕχΖ«ΑκκΉΑ±Υαά¥ΫιΒΦPTS. ΫΪ’β–©Ζ÷Ν―ΡΎΚ§κΡΤ§ΕΈ”κΨΏ”–ΒΞΗωΑκκΉΑ±ΥαΒΡΕΧκΡ–ρΝ–Θ®≥ΤΈΣCysTagΘ©»ΎΚœ±μ¥οΘ§ ‘ΎΗΟΤ§ΕΈ÷–“ΐ»κ”ΪΙβΆ≈ΓΔ …ζΈοΥΊΦΑΨέ““Εΰ¥ΦΘ®PEGΘ©Β»–ό ΈΚσΘ§ Ψ≠PTSΜώΒΟ»Ϊ≥ΛΒΡΡΩ±ξΒΑΑΉ. ΗΟΖΫΖ® Βœ÷ΝΥ‘ΎΚ§”–ΕύΗωΑκκΉΑ±ΥαΒΡΡΩ±ξΒΑΑΉΘ§ »γΝρ―θΜΙΒΑΑΉΘέ13ΘίΓΔ Π¬-ΡΎθΘΑΖΟΗΘέ13ΘίΓΔ »Υ…ζ≥ΛΦΛΥΊΚΆΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗTycAΘέ95ΘίΒ»ΒΑΑΉ÷ ÷–ΒΡΒΞ“ΜΑκκΉΑ±ΥαΈΜΒψΒΡ―Γ‘ώ–‘–ό Έ.
ΫϋΤΎΘ§ PlessΒ»Θέ96ΘίΫΪΗΟ≤Ώ¬‘”Π”Ο”ΎΧεΡΎΒΑΑΉ÷ –ό ΈΘ§ ‘ –μΫΪΕύ÷÷–ό ΈΘ®Αϋά®ΝΉΥαΜ·ΓΔ ““θΘΜ·ΡΘΡβΈοΚΆΖ«Χλ»ΜΑ±ΜυΥαΒ»Θ©Ά§ ±“ΐ»κΜνΒΡ’φΚΥœΗΑϊ÷–. Ά®ΙΐΧεΡΎtPTS≤Ώ¬‘Θ§ ‘ΎNaV1.5άκΉ”Ά®ΒάΒΡΑϊΡΎ”ρΚΆP2X2 ήΧεΒΡΑϊΆβ”ρ÷–“ΐ»κΝΥ–ό ΈΘ§ ―–ΨΩΝΥΖ≠“κΚσ–ό Έ‘ΎάκΉ”Ά®ΒάΙΠΡή“‘ΦΑ ήΧε”κ≈δΧεΫαΚœ÷–ΒΡΉς”Ο. ’ΐΫΜΖ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡ¥°ΝΣΒΑΑΉΖ¥ ΫΦτΫ”‘ΎΧεΆβΚΆΧεΡΎΒΡΒΑΑΉΕύΤ§ΕΈΦτΫ”ΕΦ’Ιœ÷ΝΥΝΦΚΟΒΡ–ßΙϊΘ§ ”–Άϊ‘ΎΒΑΑΉ÷ ΑκΚœ≥…÷–ΒΟΒΫΗϋΕύΒΡ”Π”Ο.
4.2ΓΓΤ§ΕΈΆ§ΈΜΥΊ±ξΦ«
‘ΎΡΩ±ξΒΑΑΉ÷ ÷–“ΐ»κΨΏ”–NMR–ß”ΠΒΡ15NΦΑ13CΒ»÷Ί‘≠Ή”ΚΥΒΡΆ§ΈΜΥΊ±ξΦ«Ω…Ω¥Ής «…œ ωΆ®ΙΐPTSΫΪΜ·―ß–ό Έ“ΐ»κΒΑΑΉ÷ ΒΡ“ΜΗωΨΏΧεάΐΉ”Θ§ ’β“ΜΦΦ θΉν÷Ί“ΣΒΡ”Π”Ο «”Ο”ΎΚΥ¥≈Ι≤’ώ≤®ΤΉ―–ΨΩά¥±μ’ςΒΑΑΉ÷ ΒΡΫαΙΙΚΆΕ·ΝΠ―ß. »ΜΕχΕ‘”ΎΫœ¥σΒΑΑΉ÷ ά¥ΥΒΘ§ »Ϊ≥ΛΆ§ΈΜΥΊ±ξΦ«ΒΡ―υΤΖ÷–œ‘÷χΒΡΖε÷ΊΒΰ ΙΒΟΚΥ¥≈Ι≤’ώΤΉΒΡΫβΈω±δΒΟάßΡ―.
Ι”ΟΖ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡPTSΩ…“‘ΫΪΨΏ”–÷ΊΆ§ΈΜΥΊ±ξΦ«ΒΡΒΑΑΉ÷ Τ§ΕΈ”κΈό±ξΦ«ΒΡΤ§ΕΈΝ§Ϋ”Τπά¥Θ§ …ζ≥…≤ΩΖ÷Τ§ΕΈ±ξΦ«ΒΡ»Ϊ≥ΛΒΑΑΉ÷ ―υΤΖΘ§ œ‘÷χΦρΜ·ΚΥ¥≈Ι≤’ώ≤®ΤΉ. NakamuraΒ»Θέ46Θί”Ύ1998Ρξ Ι”ΟΝΥά¥Ή‘Ι≈ΨζPyrococcus furiosusΒΡΡΎΚ§κΡΘ®PI-PfuIΘ©Θ§ ≤ΔΫΪΤδ»ΥΈΣΖ÷ΈΣ2ΗωΤ§ΕΈΘ§ Ζ÷±π”κΉςΈΣΚΥ¥≈Ι≤’ώ±μ’ςΒΡΑ–ΒΑΑΉRNAΨέΚœΟΗΠΝ―«ΜυΒΡCΕΥΫαΙΙ”ρΘ®ΠΝCΘ©ΒΡ2ΗωΤ§ΕΈ»ΎΚœΘ®ΆΦ15Θ©. Ψ≠Ά§ΈΜΥΊ±ξΦ«ΒΡΤ§ΕΈ”κΝμ“ΜΈ¥±ξΦ«Τ§ΕΈ‘ΎΧεΆβΜλΚœ≤ΔΖΔ…ζΖ¥ ΫΦτΫ”Ζ¥”ΠΘ§ Ή¥ΈΜώΒΟΝΥ”Ο”ΎΚΥ¥≈Ι≤’ώ±μ’ςΒΡ≤ΩΖ÷Τ§ΕΈΆ§ΈΜΥΊ±ξΦ«ΒΡΒΑΑΉ÷ ―υΤΖ.
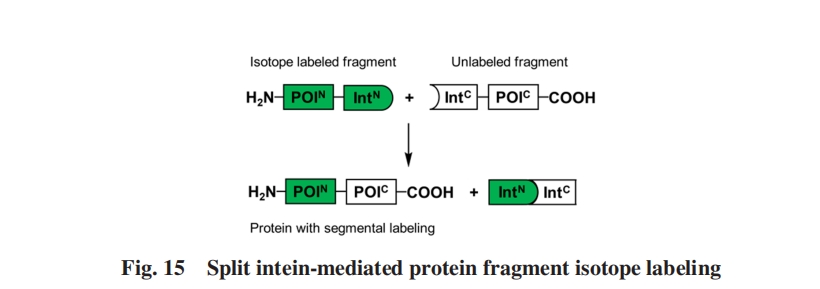
ΨΓΙήΉν≥θ±ξΦ«ΒΡΡΩ±ξΒΑΑΉ÷ Ϋω”–‘Φ9000 DaΘ§ ΒΪΗΟœΒΆ≥Κσά¥≥…ΙΠΒΊ”Ο”Ύ÷Τ±ΗΖ÷ΕΈΆ§ΈΜΥΊ±ξΦ«ΒΡ¬σ―ΩΧ«ΫαΚœΒΑΑΉΘ®42000 DaΘ©Θέ97ΘίΦΑF0F1ATPΟΗΠ¬―«ΜυΘ®52000 DaΘ©Θέ98ΘίΘ§ ÷ΛΟςΝΥΆ®ΙΐΖ÷Ν―ΡΎΚ§κΡ Βœ÷ΒΡΖ÷ΕΈΆ§ΈΜΥΊ±ξΦ«Ω…”Ο”ΎΗϋ¥σΡΩ±ξΒΑΑΉ÷ ΒΡNMRΫαΙΙΖ÷Έω.
IwaïΒ»Θέ12ΘίΩΣΖΔΝΥ“Μ÷÷Ά®Ιΐ»ΐΤ§ΕΈΒΑΑΉ÷ ΦτΫ”ά¥ Βœ÷Τ§ΕΈΆ§ΈΜΥΊ±ξΦ«ΒΡ–¬≤Ώ¬‘. άϊ”ΟΨΏ”–œύΆ§–ρΝ–ΒΪΖ÷Ν―ΈΜΒψ≤ΜΆ§ΒΡNpuDnaEΙΛ≥ΧΜ·Ζ÷Ν―ΡΎΚ§κΡΘ§ Ε‘ά¥Ή‘Lyngbya majusculaΒΡCuracin AΘ®CurAΘ©ΒΑΑΉΆ®Ιΐ»ΐΤ§ΕΈΒΑΑΉΦτΫ”ά¥ Βœ÷≤ΩΖ÷Τ§ΕΈΆ§ΈΜΥΊ±ξΦ«. ΗΟΒΑΑΉΑϋΚ§3ΗωΥ≥–ρΒΡθΘΜυ‘ΊΧεΒΑΑΉΘ®ACPΘ©ΫαΙΙ”ρ. ’β 3ΗωΫαΙΙ”ρ÷°ΦδΗΏΕ»ΒΡ–ρΝ–Ά§“Μ–‘Θ®93%~100%Θ©ΒΦ÷¬ΤδΥυ”–3ΗωΫαΙΙ”ρΨυ±ΜΆ§ΈΜΥΊ±ξΦ«ΒΡ―υΤΖΒΡNMR–≈Κ≈―œ÷Ί÷ΊΒΰ. “ρ¥ΥΘ§ ―Γ‘ώ–‘ΒΊΕ‘ΗΟ»ΐΫαΙΙ”ρΒΑΑΉ÷ ΒΡ÷––ΡΫαΙΙ”ρΫχ––ΝΥΆ§ΈΜΥΊ±ξΦ«Θ§ Ά®ΙΐΖ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡPTSΘ§ “‘‘Φ40%ΒΡ ’¬ Μώ»ΓΝΥ”Ο”ΎNMRΖ÷ΈωΒΡ≤ΩΖ÷Τ§ΕΈΆ§ΈΜΥΊ±ξΦ«ΒΡ»Ϊ≥ΛCurAΒΑΑΉ. ΗΟΖΫΖ®ΫΪ≤ΩΖ÷Τ§ΕΈΆ§ΈΜΥΊ±ξΦ«≤Ώ¬‘‘ΎΚΥ¥≈Ι≤’ώ―–ΨΩ÷–ΒΡ”Π”ΟΆΊ’ΙΒΫΝΥΕύΫαΙΙ”ρΒΑΑΉ÷–ΒΡΫαΙΙ”ρ-ΫαΙΙ”ρœύΜΞΉς”Ο―–ΨΩ.
IwaïΒ»Θέ99ΘίΜΙΩΣΖΔΝΥΫΪΆ§ΈΜΥΊ±ξΦ«ΒΡΤ§ΕΈ”κΈ¥±ξΦ«ΒΡΤ§ΕΈ‘ΎΆ§“ΜœΗΨζ≈ύ―χΈο÷–“Μ¥Έ…ζ≥…Θ§ ΥφΚσ÷±Ϋ”ΖΔ…ζΦτΫ”Ζ¥”Πά¥ΧεΡΎΜώ»ΓΖ÷ΕΈΆ§ΈΜΥΊ±ξΦ«ΒΡ»Ϊ≥ΛΒΑΑΉΒΡ≤Ώ¬‘. ΤδΜυ±Ψ‘≠άμ «‘ΎΒΞΗωœΗΑϊ≈ύ―χΙΐ≥Χ÷–ΒΡ≤ΜΆ§ ±Φδ±μ¥οΚ§Ζ÷Ν―ΡΎΚ§κΡΒΡΒΑΑΉΤ§ΕΈ«ΑΧεΘ§ ≤Δ‘ΎΗςΗω±μ¥ο≤Ϋ÷η÷°ΦδΗϋΜΜΈ¥±ξΦ«ΜρΆ§ΈΜΥΊ±ξΦ«ΧθΦΰΒΡ≈ύ―χΜυ. ’β―υΘ§ ÷Μ”–“Μ÷÷«ΑΧεΒΑΑΉΤ§ΕΈ «±ΜΆ§ΈΜΥΊ±ξΦ«ΒΡΘ§ ΦτΫ”Ζ¥”ΠΚσΒΟΒΫΒΡΦ¥ «÷Μ”–“ΜΗωΤ§ΕΈ±ΜΆ§ΈΜΥΊ±ξΦ«ΒΡΡΩ±ξΒΑΑΉΘ§ Βœ÷ΝΥΖ÷ΕΈΆ§ΈΜΥΊ±ξΦ«ΒΑΑΉ÷ ΒΡ÷±Ϋ”ΧεΡΎΜώ»ΓΘέ98Θί.
4.3ΓΓΕύκΡΚΆΒΑΑΉ÷ ΒΡΜΖΜ·
Μυ”ΎœΗΑϊΒΡΜΖκΡ…Η―ΓΖΫΖ®œύΕ‘”Ύ …ΨζΧε’Ι ΨΒ»ΧεΆβ…Η―ΓΖΫΖ®Θ§ Ω…“‘ΗϋΈΣΈ»ΫΓΒΊΒΟΒΫΨΏ”–ΧεΡΎΜν–‘ΒΡΜΖκΡΘ§ ≤ΔΦφΨΏΫœΒΆΒΡœΗΑϊΕΨ–‘ΓΔ ΫœΚΟΒΡ»ήΫβΕ»Β»–‘÷ . Ζ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡPTS“ρΤδΤ§ΕΈΩ…“‘÷±Ϋ”Ά®Ιΐ±μ¥οΜώ»Γ≤Δ«“Ω…“‘ΫΪΝΫ≤ύΒΡΕύκΡΤ§ΕΈΝ§Ϋ”Τπά¥ΒΡΧΊ–‘Θ§ Ω…±Μ”Οά¥‘ΎœΗΑϊΡΎ…ζ≥…ΜΖκΡ. Φ¥Ά®Ιΐ2ΗωΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΒΡΫαΚœΘ§ Τ» ΙΤδΝΫ≤ύΝ§Ϋ”ΒΡœΏ–‘ΕύκΡΒΡNΕΥΚΆCΕΥΆ®ΙΐκΡΦϋΆΖ-Έ≤Ν§Ϋ”Θ§ Άξ≥…ΜΖΜ·Θ®ΆΦ16Θ©.

ΈΣ¥ΥΘ§ BenkovicΒ»Θέ2ΘίΩΣΖΔΝΥΖ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡΕύκΡΚΆΒΑΑΉ÷ ΜΖΉ¥Ν§Ϋ”Θ®SICLOPPSΘ©ΦΦ θ. ’βœνΦΦ θΩ…“‘ Βœ÷ΧεΡΎΒΡΜΖκΡΈΡΩβΙΙΫ®Θ§ ≤ΔΫαΚœΙΠΡή…Η―ΓΘ§ ΒΟΒΫΝΥ–μΕύΨΏ”–…ζΈοΜν–‘ΒΡΜΖκΡΘ§ »γά“Α±ΥαΟΗ“÷÷ΤΦΝpseudostellarin FΘέ2ΘίΦΑΦΉΜυΉΣ“ΤΟΗ“÷÷ΤΦΝΒ»Θέ100Θί.
ΈΣΝΥ‘ωΦ”…ζ≥…ΜΖκΡΒΡΙΠΡήΜυΆ≈Εύ―υ–‘Θ§ SchultzΒ»Θέ101ΘίΆ®Ιΐ”κ“≈¥ΪΟή¬κά©’ΙΦΦ θΫΪΖ«Χλ»ΜΑ±ΜυΥα≤ε»κΒΫœΒΆ≥…ζ≥…ΒΡΜΖκΡ÷–Θ§ ΖαΗΜΝΥΩ……ζ≥…ΜΖκΡΈΡΩβΒΡΕύ―υ–‘Θ§ «“…Η―Γ≥ωΝΥHIVΒΑΑΉΟΗΜΖκΡ“÷÷ΤΦΝ. LindquistΒ»Θέ102ΘίΙΙΫ®ΝΥΒΎ“ΜΗωΫΆΡΗΦφ»ίΒΡΜΖκΡΈΡΩβΘ§ ≤Δ…Η―Γ≥ωΝΥΩ…‘ΎΫΆΡΗΦΑΕ·ΈοΡΘ–Ά÷–ΧΊ“λ–‘ΫΒΒΆ ΠΝ-synucleinΕΨ–‘ΒΡΜΖκΡΖ÷Ή”.
ΈΣΝΥΩΣΖΔΩ…“‘ΒςΩΊΒΑΑΉ÷ -ΒΑΑΉ÷ œύΜΞΉς”ΟΒΡΜΖκΡΖ÷Ή”Θ§ BenkovicΒ»Θέ103Θί’ϊΚœΝΥSICLOPPS”κΖ¥œρΥΪ‘”ΫΜœΒΆ≥Θ®RTHSΘ©2÷÷ΦΦ θ. ‘ΎΥό÷ςœΗΑϊ÷–Ψ≠SICLOPPSΦΦ θ≤ζ…ζΒΡΜΖκΡΈΡΩβΆ®ΙΐRTHSœΒΆ≥÷±Ϋ”Ϋχ––…Η―ΓΘ§ ΦχΕ®≥ωΝΥΒςΫΎFKBP12-άΉ≈ΝΟΙΥΊ-FRAPœύΜΞΉς”ΟΚΆHIV-1ΒΑΑΉΟΗΚΆΚΥΧ«ΚΥή’ΥαΜΙ‘≠ΟΗœύΜΞΉς”ΟΒΡΜΖκΡΖ÷Ή”Θ§ ’Ι ΨΝΥSICLOPPSΦΦ θΙΙΫ®ΜΖκΡΈΡΩβ≤Δ”Ο”ΎΜώ»ΓΩ…ΒςΩΊΒΑΑΉ÷ -ΒΑΑΉ÷ œύΜΞΉς”ΟΒΡΜΖκΡΒΡ«±ΝΠ.
ΝμΆβΘ§ ―–ΨΩ’ΏΟ«ΜΙΩΣΖΔΝΥΫΪΖ÷Ν―ΡΎΚ§κΡ”κΝ§–χΡΎΚ§κΡΝΣ”Οά¥ΙΙΫ®ΜΖκΡΒΡΖΫΖ®Θέ104ΘίΘ§ “‘ΦΑΆ®ΙΐPTS–Έ≥…κΡΦϋΆξ≥…κΡΜΖΜ·ΚσΘ§ ‘Όάϊ”ΟΑκκΉΑ±Υα≤–Μυ–Έ≥…¥σΜΖκΡΡΎ≤ΩΒΡΕΰΝρΦϋά¥ΙΙΫ®ΥΪΜΖκΡ≤ζΈοΒΡ≤Ώ¬‘Θέ105ΘίΘ§ ’βΧεœ÷ΝΥΖ÷Ν―ΡΎΚ§κΡœύΙΊΖΫΖ®”Ο”ΎΙΙΫ®ΜΖκΡάύΖ÷Ή”ΒΡΝΦΚΟ–ßΙϊΦΑΙψΖΚ”Π”Ο.
Μυ”ΎΖ÷Ν―ΡΎΚ§κΡΒΡΕύκΡΜΖΜ·ΜΙΩ…”κΤδΥϊ–¬–Υ…Η―ΓΦΦ θΫαΚœΘ§ ParkΒ»Θέ106ΘίΩΣΖΔΝΥ“Μ÷÷≥ΤΈΣCWCPSΘ®Custom-designed warhead-armed cyclic peptide screeningΘ©ΒΡ≤Ώ¬‘Θ§ ΫΪΩ…”κΑ–ΒΑΑΉΜν–‘ΈΜΒψΫαΚœΒΡΒ·ΆΖΜ·―ßΜυΆ≈“ΐ»κ”…Ζ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡPTS…ζ≥…ΒΡΜΖκΡ“÷÷ΤΦΝΘ§ ‘ΌΆ®ΙΐΫΆΡΗΥΪ‘”ΫΜΫχ––œΗΑϊΡΎ…Η―Γ. ΙΙœσΈ»Ε®ΒΡΜΖκΡ÷–ΒΡΒ·ΆΖΜ·―ßΜυΆ≈”κΜν–‘ΈΜΒψΫαΚœΚσΒΡΒΡΫβάκΗϋ¬ΐΘ§ «ΉΚΆΝΠœ‘÷χ‘ω«Ω. Ά®ΙΐΗΟΖΫΖ®ΖΔœ÷ΝΥ“Μ÷÷’κΕ‘Α©÷ΔΑ–ΒψHDAC8”––ßΒΡ“÷÷ΤΦΝCY5-6QΘ§ Τδ±μœ÷≥ωΚή«ΩΒΡΫαΚœ«ΉΚΆΝΠΘ®KD=15.1 nmol/LΘ©ΚΆ“÷÷ΤΉς”ΟΘ®IC50=0.61 ΠΧmol/LΘ©. ’β±μΟςΜυ”ΎΖ÷Ν―ΡΎΚ§κΡΒΡΖΫΖ® «œΗΑϊΡΎΙΙΫ®ΜΖκΡΒΡ“ΜΗωœύΕ‘Έ»ΫΓΒΡ≤Ώ¬‘Θ§ ‘Ύ”–«ΑΆΨΒΡΜΖκΡ“©Έο«ΑΧεΒΡΖΔœ÷÷–ΖΔΜ”Ής”Ο.
4.4ΓΓΧθΦΰ–‘ΒΑΑΉΦτΫ”
ΧθΦΰ–‘ΒΑΑΉΦτΫ”Θ®CPSΘ©Φ¥Ά®Ιΐ“ΜΗωΆβ‘¥ΒςΫΎΤςά¥ΦΛΜνΜρ“÷÷ΤΒΑΑΉΦτΫ”.Μυ”ΎΖ÷Ν―ΡΎΚ§κΡΒΡCPSœΒΆ≥ΒΡΜυ±Ψ‘≠άμ «ΫΪ»ΥΙΛ…ηΦΤΒΡ≤ΜΡήΉ‘ΖΔΫαΚœΒΡΖ÷Ν―ΡΎΚ§κΡ»ΎΚœΒΫΡΩ±ξΒΑΑΉΒΡ2ΗωΤ§ΕΈ÷–Θ§ Ά®Ιΐ“ΐ»κΧΊΕ®Άβ‘¥Ζ÷Ή”ΜρΙβ’’Β»ΖΫ Ϋ÷ΊΫ®ΤδΦτΫ”Μν–‘ΘΜΜρ «‘ΎΜν–‘ΈΜΒψ≤–Μυ¥ΠΜρΗΫΫϋΫχ––Μ·―ß–ό ΈΘ§ ΥφΚσΆ®ΙΐΙβ”’ΒΦΓΔ ΟΗ«–Β»ΖΫ Ϋ»Ξ≥ΐ–ό ΈΘ§ ¥”Εχ ΙΤδΜ÷Η¥ΦτΫ”Μν–‘. “ρ¥ΥΘ§ Ω…“‘Ά®ΙΐΩΊ÷ΤΡΎΚ§κΡΒΡΜν–‘Θ§ ά¥ΒΟΒΫ“Μ÷÷Υφ“βΓΑΦΛΜνΓ±»ΈΚΈΒΑΑΉ÷ ΒΡΖΫΖ®Θ§ …θ÷Ν‘ΎΧεΡΎ“≤Ω…“‘ ”Ο.
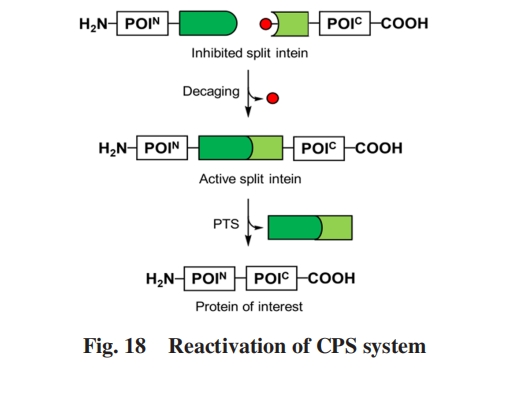
»ΥΙΛΖ÷Ν―ΒΡSce VMA ΡΎΚ§κΡ‘ΎIntNΚΆ IntCΤ§ΕΈ÷°ΦδΫω±μœ÷≥ωΖ«≥ΘΒΆΒΡ«ΉΚΆΝΠΘ§ ‘ΎΒΑΑΉ÷ ΦτΫ”÷–ΦΗΚθΈόΜν–‘. MuirΒ»Θέ107ΘίΫΪάΉ≈ΝΟΙΥΊΫαΚœΫαΙΙ”ρFKBP12ΚΆFRB»ΎΚœΒΫ2ΗωΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΚσΘ§ Φ”»κάΉ≈ΝΟΙΥΊΘ§ Ω…”’ΒΦ2ΗωΤ§ΕΈœύΜΞΩΩΫϋ≤ΔΫαΚœΘ§ ΫχΕχΒΦ÷¬Ζ÷Ν―ΡΎΚ§κΡΤ§ΕΈœύΜΞΩΩΫϋ≤Δ’έΒΰ≥…ΨΏ”–PTSΜν–‘ΒΡΗ¥ΚœΈοΘέΆΦ17Θ®AΘ©Θί. Ά®Ιΐ Βœ÷ΡΘ–ΆΒΑΑΉMBPΚΆHis-tagΒΡCPSΘ§ Ή¥ΈΆξ≥…ΝΥΒΑΑΉ÷ Ζ¥ ΫΦτΫ”Ζ¥”ΠΒΡ–ΓΖ÷Ή”ΦΛΜνΒςΩΊ. MootzΒ»Θέ108Θίάϊ”ΟάύΥΤΒΡΥΦ¬ΖΘ§ ΫΪΖ÷Ν―ΡΎΚ§κΡΤ§ΕΈΖ÷±π”κFKBP12ΒΡF36MΆΜ±δΧε»ΎΚœΘ§ ΗΟΆΜ±δΧεΈό–ηΆβΦ”–ΓΖ÷Ή”Φ¥Ω…ΖΔ…ζΆ§‘¥ΕΰΨέΜ·Θ§ ΫχΕχΫιΒΦΖ÷Ν―ΡΎΚ§κΡΖΔ…ζPTS. Εχ¥Υ ±‘ΌΦ”»κάΉ≈ΝΟΙΥΊΒ»–ΓΖ÷Ή”Θ§ ΜαΆ®Ιΐ”κΗΟΫαΙΙ”ρΫαΚœΕχΉηΕœΤδΕΰΨέΜ·Θ§ ΫχΕχ Βœ÷PTSΖ¥”ΠΒΡΧθΦΰ–‘“÷÷ΤΘ§ ΧαΙ©ΝΥ“Μ÷÷«±‘ΎΒΡΫœΈΣΆ®”ΟΒΡ ÷ΕΈ”Ο”ΎΖ≠“κΚσΒΡΒΑΑΉ÷ ΙΠΡήΒςΩΊ. ≥ΐ–ΓΖ÷Ή”œλ”ΠΒΡCPSœΒΆ≥ΆβΘ§ MuirΒ»Θέ109ΘίΜΙΩΣΖΔΝΥΙβœλ”Π–‘CPSΧεœΒΘ§ ΫΪ‘¥”ΎΡβΡœΫφΒΡΜα‘ΎΙβ’’œ¬ΖΔ…ζΕΰΨέΒΡΙβΟτ…ΪΥΊBΘ®PhyBΘ©”κΙβΟτ…ΪΥΊœύΜΞΉς”Ο“ρΉ”3Θ®PIF3Θ©”κSce VMAΖ÷Ν―ΡΎΚ§κΡΝΫΕΥΤ§ΕΈΖ÷±π»ΎΚœΘέΆΦ17Θ®BΘ©Θί. άύΥΤ”ΎFKBP12”κFRBΕ‘άΉ≈ΝΟΙΥΊΒΡœλ”ΠΘ§ PhyB”κPIF3‘ΎΙβ’’œ¬ΖΔ…ζΕΰΨέΘ§ ΙΒΟVMAΖ÷Ν―ΡΎΚ§κΡ“≤œύΜΞΩΩΫϋ≤ΔΜ÷Η¥ΦτΫ”Μν–‘. άϊ”Ο’β“ΜΧεœΒΘ§ “‘MBP-Flag tagΉςΈΣΡΘ–ΆΦτΫ”≤ζΈο‘ΎΫΆΡΗ÷– Βœ÷ΝΥΙβœλ”Π–‘CPS. ¥ΥΆβΘ§ SilverΒ»Θέ110ΘίΜΙΫΪ»ΥΙΛ…ηΦΤΒΡΜΞœύ÷°ΦδΨΏ”–Ϋœ«Ω«ΉΚΆΝΠΒΡΨμ«ζ¬ί–ΐΘ®Coiled coilΘ§ CCΘ©“ΐ»κVMAΖ÷Ν―ΡΎΚ§κΡΘ§ Βœ÷ΝΥ”ΪΙβΥΊΟΗΒΡCPS.

…œ ωCPSœΒΆ≥ΕΦ «Ά®ΙΐΫΪΨΏ”–‘Ύ“ΜΕ®ΧθΦΰœ¬ΨΏ”–ΫœΗΏΫαΚœΝΠΒΡΆβΦ”ΫαΙΙ”ρ”κ±Ψ…μΫαΚœΝΠΫœΒΆΒΡVMAΖ÷Ν―ΡΎΚ§κΡ»ΎΚœ Βœ÷ΒΡ. ΕχMuirΒ»Θέ111Θί±®ΒάΝΥΆ®Ιΐ“ΐ»κΤδΥϋΜυΆ≈Θ§ “÷÷Τ±Ψ…μΨΆΨΏ”–ΫœΗΏΫαΚœΝΠΒΡΖ÷Ν―ΡΎΚ§κΡΒΡΜν–‘Θ§ ‘Ό‘Ύ“ΜΕ®ΧθΦΰœ¬“Τ≥ΐ“÷÷ΤΜυΆ≈Άξ≥…Ά―ΝΐΘ§ ¥”Εχ Βœ÷CPSΒΡ–¬≤Ώ¬‘Θ®ΆΦ18Θ©. Κœ≥…ΝΥ‘ΎPTSΒΡΙΊΦϋΥΩΑ±Υα≤–Μυ…œΆ®ΙΐΠ¬-τ«Μυ–Έ≥…ΒΡθΞΦϋΕχΖ«≥ΘΙφΒΡΆ®ΙΐΠΝ-Α±ΜυΒΡθΘΑΖΦϋΝ§Ϋ”Κσ–χΤ§ΕΈΒΡSsp DnaEΡΎΚ§κΡΤ§ΕΈάύΥΤΈοΘ§ «“ΤδΠΝ-Α±ΜυΜυΆ≈±ΜΆβΦ”ΒΡ±ΘΜΛΜυΆ≈–ό Έ. ’βΉηΕœΝΥΗΟΤ§ΕΈΒΡO-NθΘΜυΉΣ“ΤΙΐ≥ΧΘ§ ΙΒΟΦτΫ”Ζ¥”ΠΈόΖ®ΖΔ…ζ. ±ΘΜΛΜυΆ≈Ω…“‘Ά®ΙΐΉœΆβœΏ’’…δΜρΒΑΑΉΟΗΦτ«–»Ξ≥ΐΘ§ ¥”ΕχΜ÷Η¥ΗΟΤ§ΕΈΒΡΦτΫ”Ζ¥”ΠΜν–‘. Ά®Ιΐ÷Τ±ΗΩΙΨζκΡmagaininΒΡάύΥΤΈο’Ι ΨΝΥΗΟCPSœΒΆ≥ΒΡ«±‘Ύ”Π”Ο. MootzΒ»Θέ112ΘίΫΪΤυΫώ“―÷ΣΦτΫ”Ζ¥”ΠΥΌ¬ ΉνΩλΒΡgp41-1Ζ÷Ν―ΡΎΚ§κΡNΕΥΤ§ΕΈ÷–±Θ ΊΒΡ±Ϋ±ϊΑ±ΥαΆ®Ιΐ“≈¥ΪΟή¬κά©’ΙΦΦ θΆΜ±δΈΣΩ…ΙβΜνΜ·ΒΡΝΎœθΜυή–Μυά“Α±ΥαΘέO-Θ®2-NitrobenzylΘ©-L-tyrosineΘ§ ONBYΘί. ‘Ύ365 nmΒΡΉœΆβœΏ’’…δœ¬Θ§ ΝΎœθΜυή–Μυ±ΘΜΛΜυ±ΜΆ―≥ΐΘ§ ΦτΫ”Ζ¥”Π“‘”κ“Α…ζ–Άgp41-1Ζ÷Ν―ΡΎΚ§κΡΦΗΚθœύΆ§ΒΡΥΌ¬ ΖΔ…ζ. ‘Ϋά¥‘ΫΕύΒΡCPS≤Ώ¬‘ΒΡ≥ωœ÷Θ§ ΙΒΟΗΟΖΫΖ®”–Άϊ≥…ΈΣΩ…“‘‘Ύ ±Ω’≤ψΟφ…œΒςΩΊΧεΡΎΒΑΑΉ÷ ΙΠΡήΒΡ ÷ΕΈ.
5 ΉήΫα”κ’ΙΆϊ
Ψ≠Ιΐ―–ΨΩ’ΏΒΡ≥÷–χΧΫΥςΘ§ Ζ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡΒΑΑΉ÷ Ζ¥ ΫΦτΫ”“―Ψ≠”…“Μ÷÷”–»ΛΒΡΒΑΑΉ÷ Ζ≠“κΚσœ÷œσΖΔ’Ι≥…ΈΣ“Μ÷÷”Π”ΟΙψΖΚΒΡΗΏ–ßΝ§Ϋ”ΖΫΖ®. Ε‘ΒΑΑΉ÷ Ζ¥ ΫΦτΫ”ΒΡΜ·―ßΙΐ≥ΧΒΡΫβΈωΘ§ Ι»ΥΟ«ΗϋΚΟΒΊάμΫβΖ÷Ν―ΡΎΚ§κΡΫαΙΙ”κΙΠΡή÷°ΦδΒΡΙΊœΒ. ΩλΥΌΖ¥ ΫΦτΫ”Ζ÷Ν―ΡΎΚ§κΡΒΡΖΔœ÷ΓΔ ΡΎΚ§κΡΒΡ–ρΝ–ΫχΜ·ΓΔ ΡΎΚ§κΡΈόΚέΦτΫ”≤Ώ¬‘ΒΡΩΣΖΔΓΔ ’ΐΫΜΡΎΚ§κΡΩβΒΡΫ®ΝΔΒ»Θ§ ΕΦ¥σ¥σΗΡΫχΝΥΜυ”ΎΡΎΚ§κΡΒΡΝ§Ϋ”ΦΦ θΘ§ ΙΤδ≥…ΈΣ»’ΫΞ≥… λΒΡΒΑΑΉ÷ Ν§Ϋ”ΙΛΨΏΘ§ Ζ÷Ν―ΡΎΚ§κΡΫιΒΦΒΡΒΑΑΉ÷ Ζ¥ ΫΦτΫ”ΦΦ θΒΡ”Π”Ο“―Ψ≠≈ν≤ΣΖΔ’Ι. Ζ÷Ν―ΡΎΚ§κΡœΒΆ≥¥ΌΫχΝΥΫαΙΙ…ζΈο―ßΒΡΖΔ’ΙΘ§ ΧΊ±π «ΚΥ¥≈Ι≤’ώ≤®ΤΉΫαΙΙΫβΈω÷–ΒΡ”Π”ΟΘΜΖ÷Ν―ΡΎΚ§κΡœΒΆ≥“≤¥ΌΫχΝΥΚœ≥…ΜρΑκΚœ≥…Ηϋ¥σΓΔ ΗϋΗ¥‘”ΓΔ ¥χ”–Εύ÷÷–ό ΈΒΡΒΑΑΉ÷ ΘΜ¥ΥΆβΘ§ Μυ”ΎΖ÷Ν―ΡΎΚ§κΡΦΦ θΒΡΜΖΉ¥κΡΩβ“≤“―≥…ΈΣ“©ΈοΖΔœ÷ΒΡ”––ß≤Ώ¬‘.
»ΜΕχΘ§ »ΥΟ«“≤±Ί–κ»œ ΕΒΫΡΩ«ΑΕ‘Ζ÷Ν―ΡΎΚ§κΡΒΡάμΫβΚΆ―–ΨΩ…–≤ΜΆξ…Τ. Υδ»ΜΕ‘”ΎΖ÷Ν―ΡΎΚ§κΡΫαΚœ“‘ΦΑΚσ–χΖ¥”ΠΙΐ≥Χ“―Ψ≠Ϋχ––ΝΥ―–ΨΩΘ§ ΒΪ «Ε‘”ΎΖ¥”ΠΙΐ≥Χ÷–ΗϋœΗ÷¬ΒΡΫαΙΙ±δΙΙ”κ¥ΏΜ·ΙΠΡή±δΜ·ΒΡΖ÷ΈωΜΙ±»Ϋœ¥÷≤Ύ. ΗϋΚΟΒΊάμΫβΝρθΞΫΜΜΜΙΐ≥Χ÷–2ΗωέœΜυ»γΚΈΩΩΫϋΘ§ ÷ßΝ¥ΝρθΞ»γΚΈΕ‘ΧλΕ§θΘΑΖΜΖΜ·Ιΐ≥Χ≤ζ…ζ¥ΌΫχΘ§ ΕΦ”–÷ζ”ΎΗϋΚΟΒΊ–≠ΒςΦτΫ”Ζ¥”ΠΒΡΗςΗω≤Ϋ÷ηΒΡΕ·ΝΠ―ßΘ§ ¥”ΕχΧαΗΏ’ϊΧεΒΡΝ§Ϋ”–߬ Θ§ ±ήΟβC-ΕΥΜρN-ΕΥΥ°ΫβΗ±≤ζΈοΒΡ≥ωœ÷. ¥ΥΆβΘ§ ΡΩ«ΑΕ‘Ζ÷Ν―ΡΎΚ§κΡ‘Ύ«Ω±δ–‘ΓΔ ΗΏΈ¬ΓΔ ―θΜ·–‘Β»ΧθΦΰœ¬ΒΡΈ»ΫΓ–‘―–ΨΩ…–≤Μ≥δΉψΘ§ Ω…Ρή‘Ύ”κΤδΥϋΜ·―ßΖ¥”ΠΝΣ”Ο ±¥χά¥≤Μ»ΖΕ®–‘.
ΥφΉ≈ΗϋΕύΒΡΈΔ…ζΈοΜυ“ρΉι±Μ≤β–ρΘ§ Ζ÷Ν―ΡΎΚ§κΡΒΡ ΐΝΩΫΪ―ΗΥΌ‘ω≥ΛΘ§ Έ¥ά¥”–ΆϊΖΔœ÷ΨΏ”–ΕάΧΊ–‘÷ Θ®άΐ»γΡήΙΜΡΆ ήΗΏΈ¬ΓΔ ΗΏ―ΈΒ»ΦΪΕΥΧθΦΰΘ©ΒΡ–¬–ΆΖ÷Ν―ΡΎΚ§κΡΘ§ ΖαΗΜ»ΥάύΕ‘Ζ÷Ν―ΡΎΚ§κΡΒΡ»œ ΕΘ§ Ϋ“ Ψœ÷Ϋώ…–Έ¥»œ ΕΒΫΒΡ«±ΝΠΘ§ ΈΣΖ÷Ν―ΡΎΚ§κΡΦΦ θΒΡΖΔ’Ι¥χά¥–¬ΒΡΜζ”ωΘ§ ≤ΔΉν÷’ΜώΒΟΆ®”Ο–ΆΡΎΚ§κΡΘ§ ΡήΙΜΕ‘»ΈΚΈΆβœ‘κΡ–ρΝ–ΨυΨΏ”–ΦτΩλΥΌΫ”Ε·ΝΠ―ßΘ§ ≤ζ¬ ΗΏΘ§ Ζ÷Ν―Τ§ΕΈ“Ή”ΎΆ®ΙΐΜ·―ßΚœ≥…ΦΦ θΜώ»ΓΘ§ «“Ν§Ϋ”Μν–‘“Ή”ΎΆ®ΙΐΆβ‘¥¥ΧΦΛΩΊ÷ΤΒ».
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
