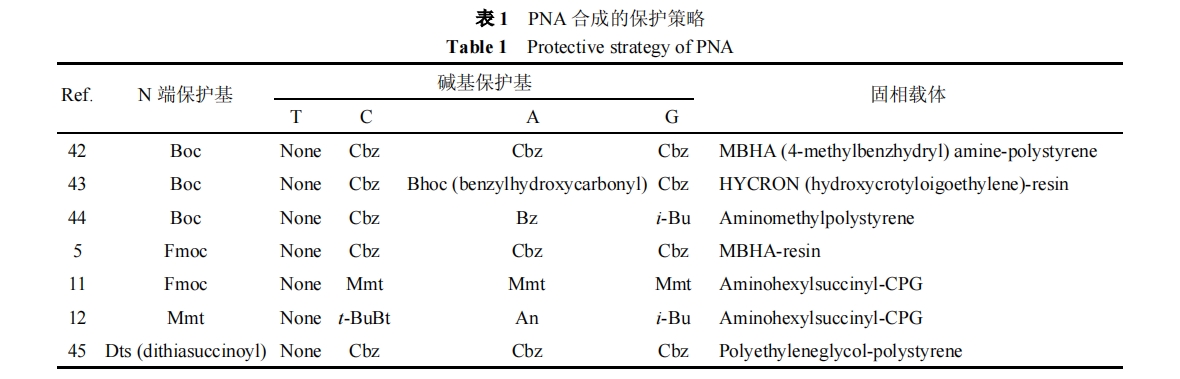
’Σ“Σ κΡΚΥΥα «Ι―ΚΥή’ΥαΡΘΡβΈο, ΥϋΒΡΧ«-ΝΉΥαΙ«Φή±Μ N-(2-Α±Μυ““Μυ)Η Α±ΥαΙ«ΦήΥυ¥ζΧφ. ΈΣΝΥ”≈Μ·κΡΚΥΥαΒΡ–‘÷ , Ης÷÷ΫαΙΙΒΡκΡΚΥΥα(PNA)ΒΞΧε±ΜΚœ≥…≥ωά¥. Ήέ ωΝΥκΡΚΥΥαΒΡΚœ≥…ΓΔΫαΙΙ–ό ΈΦΑ”Π”Ο.
Ι―ΚΥή’Υα‘Ύ÷ΈΝΤ”Ο“©ΈοΓΔΖ÷Ή”…ζΈο―ß―–ΨΩΙΛΨΏΦΑ’ο Εœ ‘ΦΝ…œ”Π”ΟΙψΖΚ. Ι―ΚΥή’Υα”Π”ΟΒΡ÷ς“Σ÷Τ‘Φ‘Ύ”ΎΥϋ»ί “Ή±ΜΚΥΥαΟΗΥυΫΒΫβΓΔ‘”ΫΜΚσ»»Έ»Ε®–‘≤ν“‘ΦΑΜα”κΒΑΑΉΒ» Ζ«ΧΊ“λ–‘ΫαΚœΒ» . ΈΣΗΡ‘λΙ―ΚΥή’ΥαΒΡάμΜ·ΚΆ…ζΈο―ßΧΊ –‘, –μΕύΙ―ΚΥή’ΥαάύΥΤΈο±Μ―–ΨΩ≥ωά¥. κΡΚΥΥα «Ι―ΚΥή’ ΥαΡΘΡβΈο÷–ΒΡΫœΈΣ÷Ί“ΣΒΡ“Μ÷÷.
1991 Ρξ Nielsen Β»[1]±®ΒάΝΥ”Ο N-(2-Α±Μυ““Μυ)Η Α± ΥαΙ«Φή¥ζΧφΧ«-ΝΉΥαθΞΙ«ΦήΉςΈΣ÷ΊΗ¥ΫαΙΙΒΞ‘Σ(ΆΦ 1), Κœ ≥…ΝΥ“‘κΡΦϋΝ§Ϋ”ΒΡΙ―ΚΥή’ΥαΡΘΡβΈο , ≥ΤΈΣκΡΚΥΥα (Peptide nucleic acid, PNA). ΨΓΙή PNA ‘ΎΫαΙΙ…œœύΕ‘Ι― ΚΥή’Υα”–ΝΥœ‘÷χΒΡΗΡ±δ, ΒΪ PNA ”κΜΞ≤ΙΚΥΥα÷°ΦδΒΡΫα Κœ»‘Ήώ―≠ΦνΜυΜΞ≤Ι≈δΕ‘‘≠‘ρ[2], …θ÷Ν±»Χλ»ΜΚΥή’ΥαΨΏ”– ΗϋΗΏΒΡ«ΉΚΆ–‘[3], ”–Ϋœ«ΩΒΡΩΙΚΥΥαΟΗΚΆΒΑΑΉΥ°ΫβΟΗΫΒΫβ ΒΡΡήΝΠ[4] . ΩΤ―–»Υ‘±…ηΦΤΚœ≥…ΝΥΗς÷÷ΫαΙΙΒΡ PNA, “‘”≈ Μ·Τδ…ζΈο―ßΈ»Ε®–‘ΚΆάϊ”ΟΕ»ΓΔΑ–ΫαΚœΧΊ–‘“‘ΦΑ“©¥ζΕ·ΝΠ ―ßΧΊ–‘, ≤ΔΫΪ PNA ”Π”Ο”Ύ’οΕœΚΆ÷ΈΝΤΒ»ΖΫΟφ, ¥”ΕχΩΣ¥¥ ΝΥ“ΜΗω–¬ΒΡ―–ΨΩΝλ”ρ. ±ΨΈΡΕ‘ PNA ΒΡΚœ≥…ΓΔ–ό ΈΚΆ”Π”ΟΫχ––Ήέ ω.
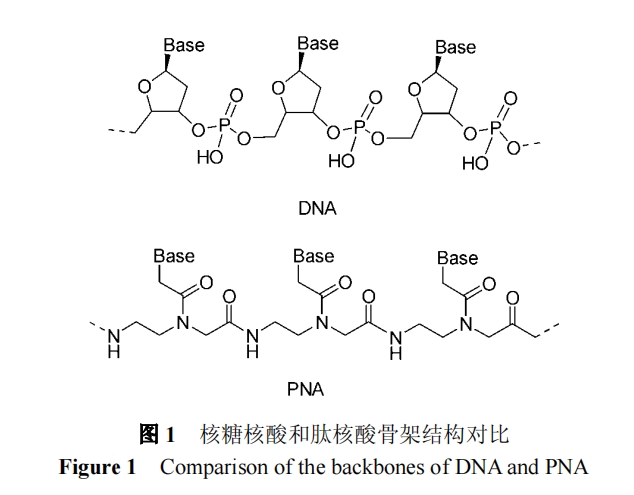
1 PNA ΒΞΧεΒΡΚœ≥…
Ης÷÷Ης―υΒΡΙΙΫ®ΡΘΩι±Μ”Ο”ΎΚœ≥… PNA ΦΑΤδάύΥΤΈο,Αϋά®Ι«ΦήΫαΙΙΓΔ‘Ύ N-(2-Α±Μυ““Μυ)Η Α±Υα…œΝ§Ϋ” ÷–‘ΚΆ Ζ« ÷–‘ΒΡΜυΆ≈ΓΔΦνΜυΒΡάύ–ΆΒ».
1.1 Ψ≠ΒδΒΡ PNA Ι«ΦήΚœ≥…
Ψ≠ΒδΒΡ PNA ΒΡΙ«ΦήΒΞΧε « N-(2-Α±Μυ““Μυ)Η Α±Υα, ‘ΎΗ Α±ΥαΒΡΒΣ…œΝ§Ϋ”ΦνΜυΒΡ―ή…ζΈο. Ά®≥Θœ»Κœ≥…ΕΥΜυ N ”–±ΘΜΛΜυΒΡΑ±Μυ““ΜυΗ Α±ΥαθΞ, »ΜΚσ‘ΌΫΪΦνΜυ―ή…ζΈοΝ§ ‘ΎΈ¥ ή±ΘΜΛΒΡΒΣ…œ. ≥Θ”ΟΒΡΖΫΖ®”–:
1.1.1 ΆιΜυΜ·Ζ¥”Π
“‘““ΕΰΑΖΜρΑ±Μυ““κφΈΣ‘≠Νœ, ”ꬱ¥ζ““Υα―ή…ζΈοΫχ ––ΆιΜυΜ·Ζ¥”Π, ”ΟΒΡ±ΘΜΛΜυ(protect group, PG)”–: ήΧ ΦΉ―θθΘΜυ[5] (9-fluorenylmethyloxycarbonyl,Fmoc)ΓΔΕ‘ΦΉ ―θΜυ±ΫΜυΕΰ±ΫΦΉΜυ [6] (4-methoxyphenyldiphenylmethyl, Mmt)ΓΔ εΕΓ―θτ Μυ(tert-butyloxycarbonyl, Boc)[7,8] (ΆΦ 2).

1.1.2 œ·ΖρΦνΒΡΜΙ‘≠Ζ¥”Π
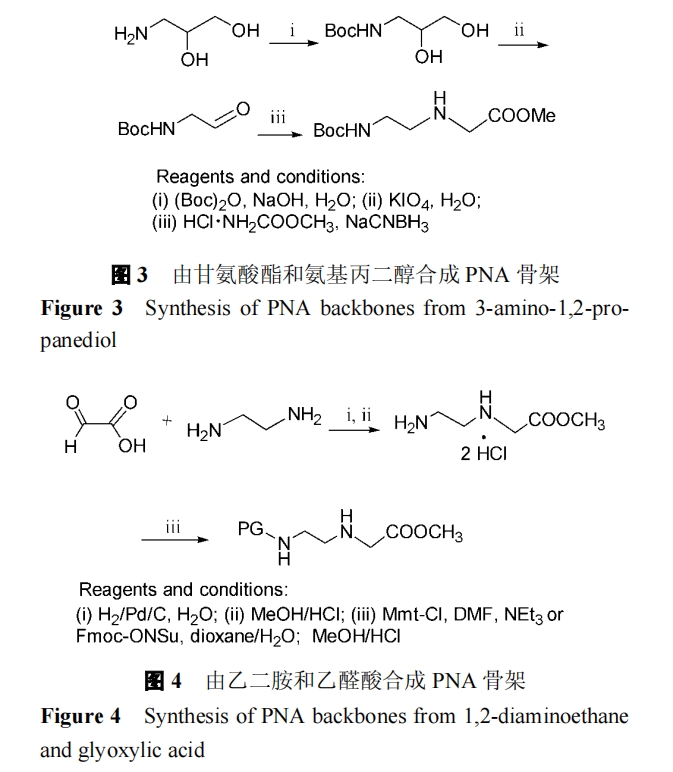
ΜΙ‘≠Η Α±ΥαθΞ”κ±ΘΜΛΒΡΑ±Μυ““»©–Έ≥…ΒΡœ·ΖρΦν[9],Υδ»Μ÷Μ ”Ο”ΎBoc±ΘΜΛΜυ, ΒΪΗΟΖΫΖ®…‘Φ”–όΗΡΦ¥Ω…”Ο”ΎΚœ≥…Ης÷÷”–≤ύΝ¥ΒΡPNAΒΞΧε[10](ΆΦ3).
ΜΙ‘≠““ΕΰΑΖ”κ““»©Υα–Έ≥…ΒΡœ·ΖρΦν, ΒΟΒΫN-(2-Α±Μυ““Μυ)Η Α±Υα, »ΜΚσ‘Ό―Γ‘ώΝ§Ϋ” Β±ΒΡ±ΘΜΛΜυ, ”–Fmoc[11]ΚΆMmt[12]Β»(ΆΦ4).
œ»ΫΪΗ Α±ΥαΜΙ‘≠≥…BocΑ±Μυ““»©, ‘Ό”κΗ Α±ΥαθΞΖ¥”Π[13](ΆΦ5).
1.1.3 Mitsunobu Ζ¥”Π
άϊ”ΟΑ±Μυ““¥Φ”κΕ‘œθΜυ±ΫΜυΦΉΜ«θΘΜυ(o-NBS)±ΘΜΛ ΒΡΗ Α±ΥαΦΉθΞΫχ–– Mitsunobu Ζ¥”Π[14](ΆΦ 6).

1.2 ‘ΎΙ«Φή…œ“ΐ»κΦνΜυ
ΥΡ÷÷ΦνΜυΕΦ «Ψ≠ΑΖΒΡΆιΜυΜ·Ζ¥”Π–Έ≥…ΦνΜυ““Υα―ή…ζΈο, ‘Ό≤…”ΟΆ®≥ΘΒΡΕύκΡΚœ≥…ΒΡΖΫΖ®, ΝΣΫαΦνΜυ““ΥαΚΆΙ«Φή…œΒΡΈ¥ ή±ΘΜΛΒΡΒΣ.
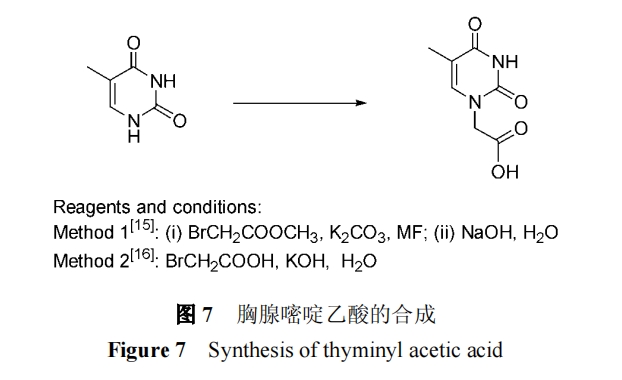
–ΊœΌύΉύΛΒΡΆιΜυΜ·Ζ¥”ΠΆ®≥Θ≤Μ–η“Σ Ι”Ο±ΘΜΛΜυΆ≈,“ρ¥ΥΒ±”κδε““ΥαθΞΖ¥”Π‘ΌΨ≠‘μΜ·[15]Μρ÷±Ϋ””κδε““ΥαΖ¥”Π[16]Φ¥Ω…ΒΟ–ΊœΌύΉύΛ““Υα(ΆΦ7).
Τδ”ύ»ΐ÷÷ΦνΜυ…œΕΦ”–ΜνΤΟΜυΆ≈, –ηœ»Φ”“‘±ΘΜΛ.ΑϊύΉύΛ…œΒΡΜνΤΟΜυΆ≈ΈΣ4ΈΜ…œΒΡΑ±Μυ, Ω…―Γ‘ώΒΡ±ΘΜΛΜυ”–: ή–―θτ Μυ(benzyloxycarbonyl,Cbz)[5]ΓΔΕ‘ εΕΓΜυ±ΫΦΉθΘΜυ(4-tert-butylbenzoyl, 4-t-BuBz)[11]ΓΔ±ΫΦΉθΘΜυ(benzoyl, Bz)[17]“‘ΦΑMmt[12]Β», ‘Ό”κδε““ΥαθΞΫχ––ΆιΜυΜ·Ζ¥”Π, »ΜΚσ‘μΜ·Φ¥ΒΟΑϊύΉύΛ““ΥαΒΡ―ή…ζΈο(ΆΦ8).
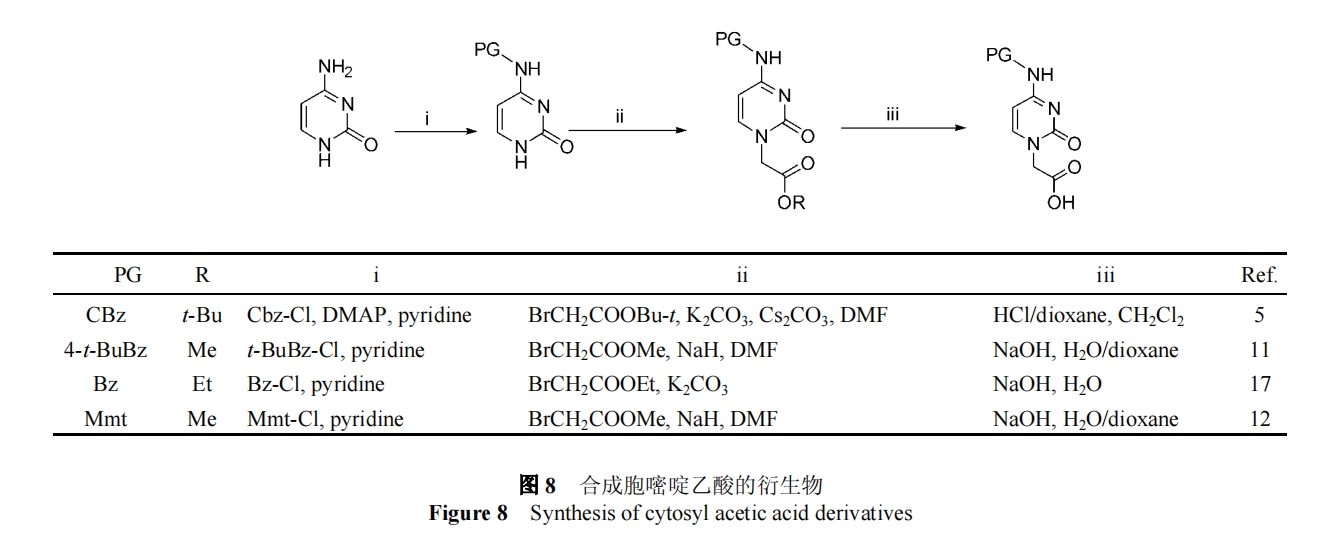
œΌύ―Ώ ΒΡ±ΘΜΛΙΐ≥Χ”κΑϊύΉύΛΜυ±ΨœύΆ§, Ω…”ΟΒΡ±ΘΜΛΜυ”– Cbz[5], Mmt[12], Ε‘ΦΉ―θΜυ±ΫΜυ[13](anisoly,An)(ΆΦ9).
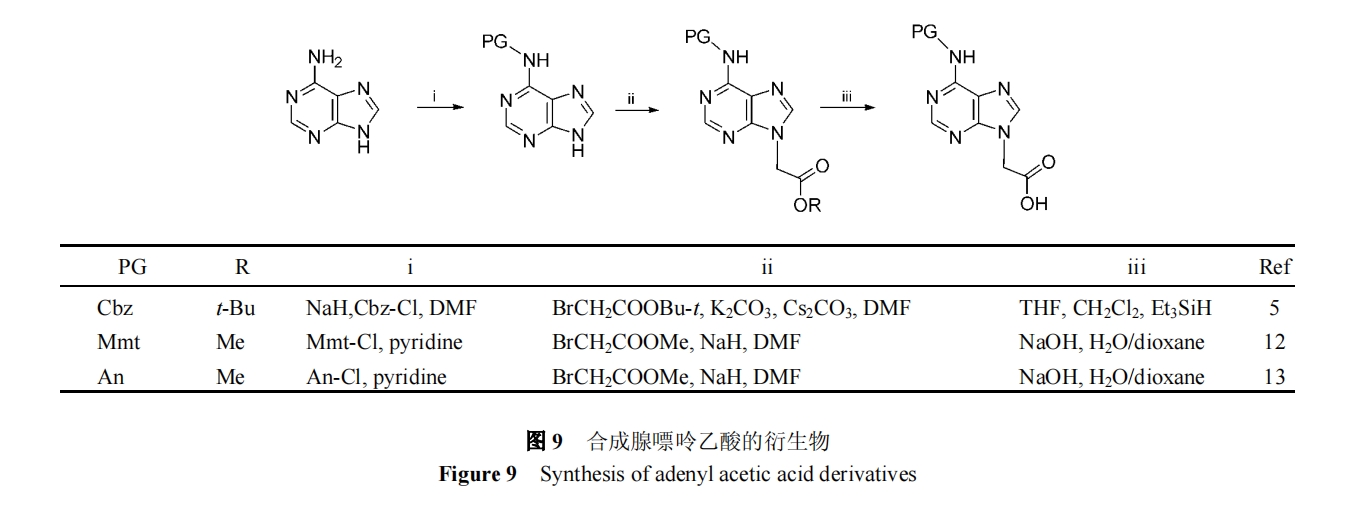
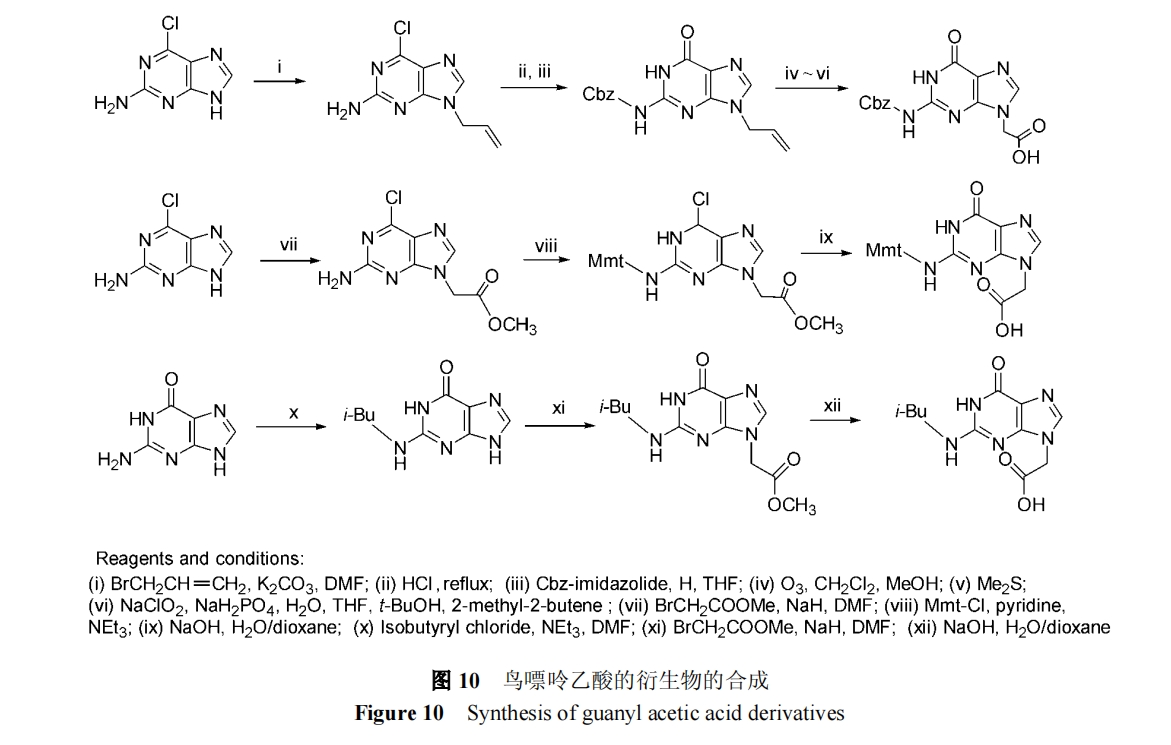
Ρώύ―Ώ ΒΡ±ΘΜΛ±»ΫœΗ¥‘”, –η“Σ‘ΎN9 ΒΡΆιΜυΜ·ΒΡΙΐ≥Χ÷–±ήΟβN7 ΆιΜυΜ·ΒΡΗ±Ζ¥”ΠΗ…»≈. “Μ÷÷≥Θ”ΟΒΡΖΫΖ® «‘ΎΆιΜυΜ·÷–”Ο2-Α±Μυ-6-¬»ύ―Ώ , ΆιΜυΜ·Κσ‘Ό‘ΎΥα–‘ΜρΦν–‘ΧθΦΰœ¬ΜΊΝς, ΫΪ¬»Υ°ΫβΉΣΜ·ΈΣτ Μυ, Ιΐ≥Χ»γΆΦ10a[5], 10b[11]. Μρ «÷±Ϋ”ΆιΜυΜ·N2Ν§”–±ΘΜΛΜυΒΡœΌύ―Ώ ,…ΪΤΉΖ÷άκN7/N9 ΝΫ÷÷ΆιΜυΜ·≤ζΈο, »ΜΚσ‘μΜ·ΒΟΡώύ―Ώ ““ΥαΒΡ―ή…ζΈο[12](ΆΦ10c).
1.3 Ε‘Ψ≠Βδ PNA ΒΡΗΡ‘λ
1.3.1 PNA Ι«ΦήΒΡ–ό Έ
ΡΩ«Α, PNA Κœ≥…ΒΡ―–ΨΩΦ·÷–‘ΎΙ«Φή–ό Έ PNA ―ή…ζ Έο…œ, «“ρΈΣ”ΟΗΡ‘λΚΆΈ¥ΗΡ‘λΒΡ PNA ΒΞΧε, ΜλΚœ÷Τ≥…Ι« ΦήΒΡΙ―ΚΥή’ΥαάύΥΤΈοΕ‘ DNA ΚΆ RNA ”–ΗϋΚΟΒΡ‘”ΫΜΧΊ –‘[18], Εχ«“Ι«Φή–ό ΈΡή”≈Μ· PNA ΒΡΧΊ–‘, »γΥ°»ή–‘ΓΔ…ζ Έοάϊ”ΟΕ»Β». ―”≥ΛΙ«ΦήΧΦΝ¥Μα Ι PNA ‘”ΫΜΜν–‘œ‘÷χΫΒ ΒΆ[19] . Β±«Α―–ΨΩΫœΕύΒΡ”–“‘œ¬ΦΗΗωΖΫΟφ.
1.3.1.1 ‘ΎΙ«Φή…œ“ΐ»κ÷ßΝ¥
“ΐ»κ÷ßΝ¥Ω… ΙΒΞΧε≥…ΈΣ ÷–‘Ζ÷Ή”, ΕχΕ‘‘”ΫΜ–‘÷ ‘ρ ”ΑœλΚή–Γ[20] . ≥Θ”ΟΦΗ÷÷“ΐ»κ≤ύΝ¥ΖΫΖ®Αϋά®Ε‘Ψ≠ΒδΒΞΧε Κœ≥…ΖΫΖ®ΒΡΗΡΫχ , άϊ”ΟΗς÷÷Χλ»Μ ΠΝ - Α±ΜυΥα “ΐ»κ÷ß Ν¥[10,20,21,14]ΓΔ≤ΜΕ‘≥Τ¥ΏΜ·«βΜ·Ζ¥”Π[22]ΓΔUgi 4CC Ζ¥”Π[23](ΆΦ 11)Β».
«Α»ΐ÷÷ΖΫΖ® «Ε‘Ψ≠Βδ PNA ΒΞΧεΚœ≥…ΖΫΖ®ΒΡΗΡΫχ. ΤδΚΟ¥Π‘Ύ”ΎΩ…“‘άϊ”ΟΗς÷÷Χλ»ΜΜρΖ«Χλ»ΜΑ±ΜυΥα‘≠Νœ, ‘≠ Νœ“ΉΒΟ, ÷ßΝ¥ΫαΙΙάύ–ΆΕύ. ΒΎΥΡ÷÷ΖΫΖ®”…”Ύ”ΟΒΫΝΥ≤ΜΕ‘ ≥Τ¥ΏΜ·«βΜ·, –η”ΟΙβ―ß¥Ω¥ΏΜ·ΦΝ, œ÷“―≤Μ‘Ό”Π”Ο. ΒΎΈε ÷÷ΖΫΖ® «“‘“λκφΓΔ τ»ΥαΓΔΑΖΓΔ»©ΜρΆΣΈΣ‘≠ΝœΒΡΕύΉιΖίΥθ ΚœΖ¥”Π. ΆΦ 11 Ηχ≥ωΒΡΫωΈΣΤδ÷–“Μάΐ, ΥΡ÷÷ΉιΖίΖ÷±πΈΣΕ‘ εΕΓΜυ -1- ΜΖΦΚœ© Μυ“λκφ (4-t-Bu-cyclohex-1-enyl iso- cyande) ΓΔN2-ή–―θτ Μυ-N9-τ»““ΜυΡώύ―Ώ (N2-Z-N2-car- boxymethylguanine)ΓΔΒΞ εΕΓ―θτ Μυ““ΕΰΑΖ(mono-Boc- ethylendiamine)ΓΔ»ΐΦΉΜυ““»©(pivalaldehyde). ¥ΥΚœ≥…ΖΫΖ®ΦρΒΞ, ≤ΔΩ…“ΐ»κΡ≥–©ΧΊ βΒΡ÷ßΝ¥, Ρή¥σ¥σά©’Ι PNA ΒΡ ÷÷άύ.

1.3.1.2 ‘ΎΙ«Φή…œ“ΐ»κΜΖΉ¥ΫαΙΙ
¥χ”–ΜΖΉ¥ΫαΙΙΒΡ PNA ”–ΚήΕύ, ΤδΚœ≥…Ιΐ≥ΧΗς“λ, Η§ Α±Υα”…”ΎΤδΧλ»ΜΫαΙΙΧΊΒψ≥…ΈΣ―–ΨΩΒΡ÷ς“Σ»»Βψ. “‘œ¬Ηχ ≥ωΦΗΗωΨΏ”–¥ζ±μ–‘ΒΡ”… 4-τ«ΜυΗ§Α±ΥαΜρΥΡ«βΏΝΩ©―ή…ζ Έο÷Τ±ΗΒΡΙ«ΦήΚ§Έε‘ΣΜΖΫαΙΙΒΡ PNA ΒΞΧεΒΡΚœ≥…ΆΨΨΕ. ΆΦ 12[24ΓΪ27] .
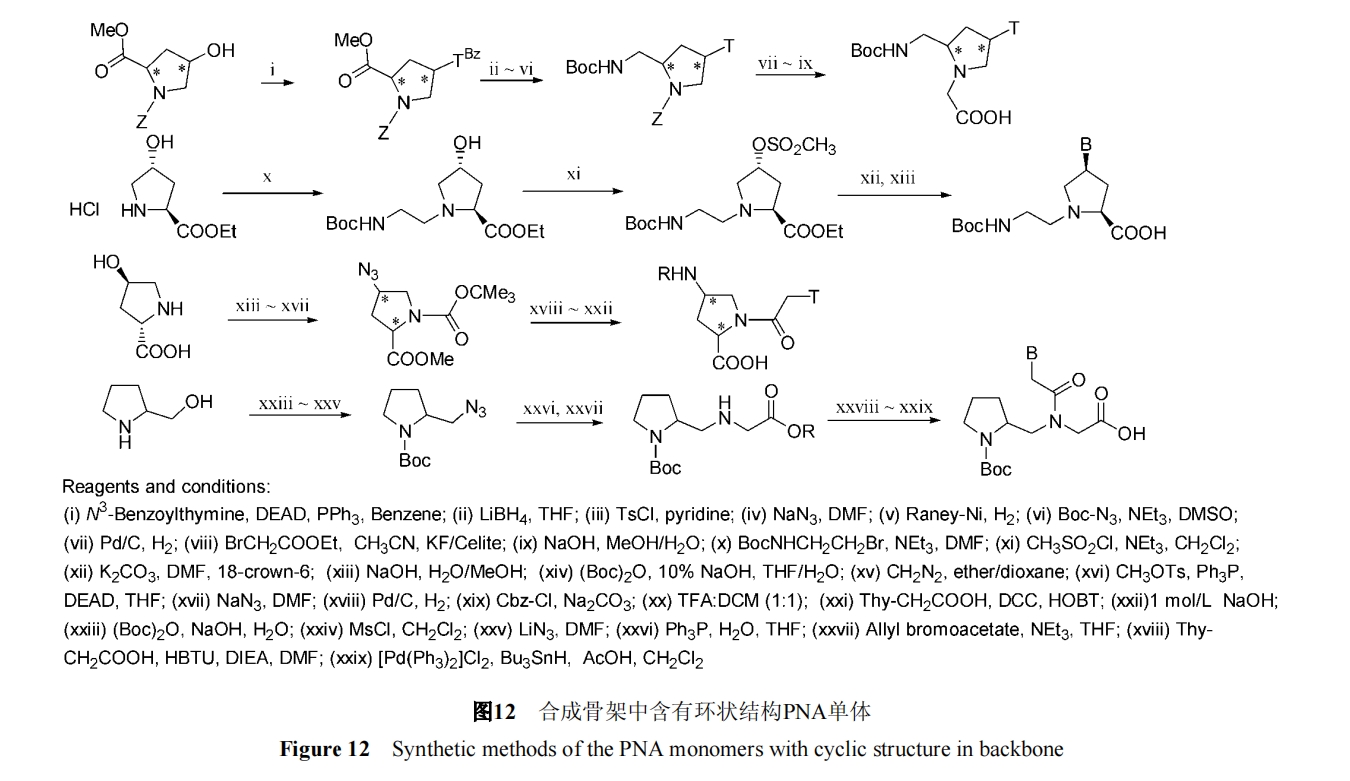
ΤδΥϋ¥χ”–ΜΖΉ¥ΫαΙΙΒΡ PNA ΒΞΧεΜΙ”–Ά§ΈΣΈε‘ΣΜΖΒΡ 1[28], 2[29], 3[30], 4[31], ΦΑΝυ‘ΣΜΖΒΡ 5[32], 6[23] “‘ΦΑ¥χ”–Νρ‘≠ Ή”ΒΡ 7, 8[33](ΆΦ 13).
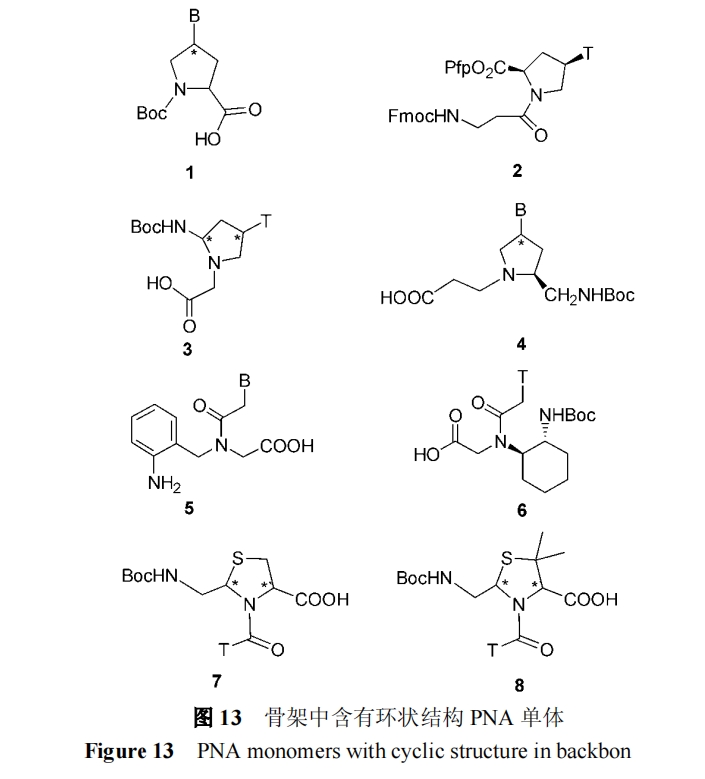
ΜΖΉ¥ΫαΙΙΒΡ“ΐ»κΕ‘ PNA ΒΡ–‘÷ ¥χά¥–μΕύ–¬ΒΡ±δΜ·. »γΕ‘ 3 ΒΡ―–ΨΩ±μΟς, ΤδΙβ―ß“λΙΙΧεΕ‘ Tm ΒΡ”Αœλ «≤Μ“Μ÷¬ ΒΡ. Κ§”–“ΜΗω(2S,4R)ΒΞΧεΒΡ PNA2 ΓΟ DNA ΒΡ Tm ÷Β±»¥ΩΨ≠ Βδ PNA ΒΞΧεΒΡ PNA2 ΓΟ DNA ΒΡ Tm ÷ΒΧαΗΏΝΥ 14 Γφ , Εχ (2S,4S)‘ρΫΒΒΆΝΥ 20 Γφ . “ρ¥Υ, Ε‘”ΎΙ«Φή÷–”–ΜΖΉ¥ΫαΙΙ ΒΡ PNA ΒΞΧε”Π≥…ΈΣΫώΚσ―–ΨΩΒΡ“ΜΗω÷ΊΒψ.
1.3.2 ΗΡ±δΦνΜυ”κΙ«ΦήΒΡΝΣΫ”ΈΜ÷Ο”κΖΫ Ϋ, ≤…”ΟΖ« θΘΑΖΦϋΒΡΖΫ ΫΫΪΦνΜυΝΣ”ΎΙ«Φή÷–ΒΡΧΦ‘≠Ή”…œ
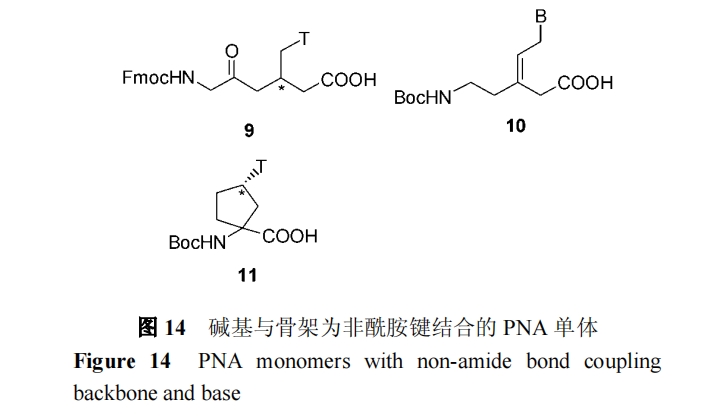
ΈΡœΉ±®Βά÷–÷ς“Σ”–“‘œ¬ΦΗ÷÷Ν§Ϋ”ΖΫ Ϋ(ΆΦ14): ΦνΜυ”κΙ«Φή“‘Έότ ΜυΒΡ÷±Ν¥CΓΣCΦϋΝ§Ϋ”9[34]; “‘œ©ΦϋΝ§Ϋ”10[35]; “‘ΜΖΉ¥ΫαΙΙΝ§Ϋ” 11[36]. ΒΪ”…”Ύ’β÷÷ΗΡ‘λΖΫ Ϋ ΙΒΟPNA ΒΡΫαΙΙ”–Κή¥σΗΡ±δ, ”–Ω…ΡήΜαΕ‘‘”ΫΜ–‘÷ ≤ζ…ζΫœ¥σΒΡΗΚΟφ”Αœλ. »γAbdel-Aziz Β»Ε‘ΒΞΧε 9 ΒΡ―–ΨΩΖΔœ÷, Ε‘”Ύ12ΨέΒΡPNAΒΆΨέΧε, ΝΫΕΥ”ΟΒΞΧε9ΧφΜΜΤδ÷–1ΓΪ3ΗωΨ≠ΒδΒΞΧε, ÷Μ ΙΤδ”κDNAΚΆRNA‘”ΫΜΒΡTm÷Β¬‘”–ΫΒΒΆ(ΓςTmΓή10 Γφ), Εχ‘ΎΒΆΨέΧεΒΡ÷–Φδ≤ε»κ“ΜΗωΒΞΧε, ΨΆΜαœ‘÷χΫΒΒΆTm÷Β. ΒΞΧε»Ϊ≤ΩΈΣ9 ΒΡΒΆΨέΧε, ‘ρΟΜ”–Ιέ≤λΒΫΤδ”κDNAΜρRNAΒΡ‘”ΫΜ.
1.3.3 Ε‘ΦνΜυΒΡΧφ¥ζ
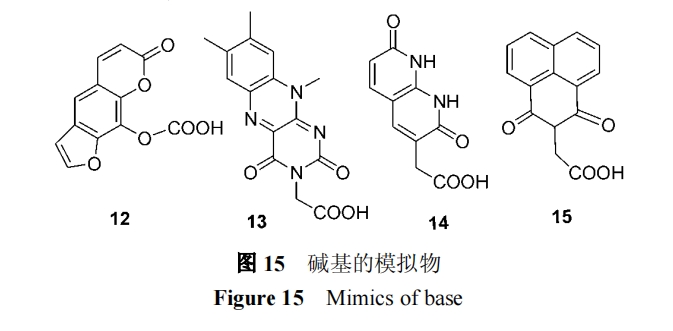
ΆΦ 15 ÷–ΒΡ 12[37], 13[38], 14[39], 15[40] “―±Μ ‘―ι”ΟΉςΦν ΜυΒΡΧφ¥ζΈο, ≤ΔΙέ≤λΒΫΝΥ”κ DNA ΒΡ‘”ΫΜ.
1.4 PNA ΒΆΨέΧεΒΡΚœ≥…
PNA÷°ΦδΒΡΝ§Ϋ”άύΥΤ”ΎΕύκΡ, “ρ¥ΥPNAΒΡΚœ≥…Ω…≤…”ΟΕύκΡΙΧœύΚœ≥…ΦΦ θ. “‘Boc/Cbz±ΘΜΛ≤Ώ¬‘ΈΣάΐ, ΤδΖ¥”ΠΙΐ≥Χ»γΆΦ16Υυ Ψ[41.
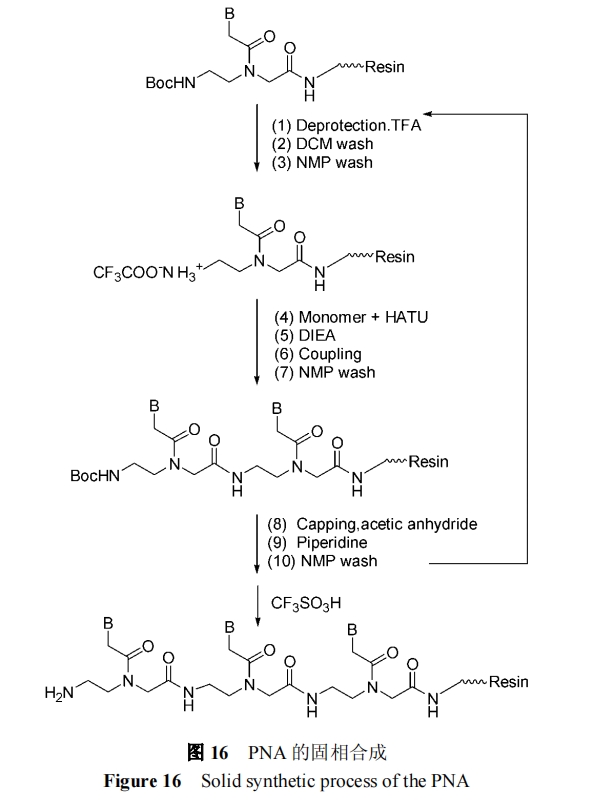
PNAΚœ≥…÷–”ΠΉΔ“β―Γ‘ώΚœ ΒΡNΕΥΚΆΦνΜυ±ΘΜΛ≤Ώ¬‘. Υδ»ΜPNAΒΞΧεΚœ≥…”ΟΒΫΒΡ±ΘΜΛΜυΚήΕύ, ΒΪ”…”Ύ ήΒΫ±ΘΜΛΚΆΆ―≥ΐΧθΦΰΦΑΙΧœύΚœ≥…ΒΡœό÷Τ, ≤Δ≤Μ «»Έ“βΝΫΗω±ΘΜΛΜυΕΦΡήΉςΈΣPNAΒΞΧεΒΡNΕΥΚΆΦνΜυ±ΘΜΛΜυΕ‘(NΕΥ/ΦνΜυ)ΒΡ±ΘΜΛ≤Ώ¬‘. ±μ 1 Ηχ≥ωΈΡœΉ÷–≥ωœ÷ΒΡ“Μ–©±ΘΜΛ≤Ώ¬‘.
2 PNA ΒΡ”Π”Ο
2.1 ÷ΈΝΤ
¥”άμ¬έ…œΩ¥, PNA”–ΖΔ’ΙΈΣΖ¥“ε“©ΈοΒΡΩ…Ρή. ÷ς“Σ“ρΈΣ:(1)PNA≤ΜΡή±ΜΚΥΥαΟΗΚΆΒΑΑΉΟΗΫΒΫβ. (2)”κDNAΚΆRNAΒΡΫαΚœΝΠ«Ω, ΧΊ“λ–‘ΗΏ.(3)PNA”κRNAΫαΚœΒΡΈ»Ε®–‘‘ΕΗΏ”Ύ”κDNAΒΡΫαΚœ.(4)PNA”κDNA–Έ≥…ΒΡPNA2/DNA»ΐ¬ί–ΐΫαΙΙΡή“ΐΤπΉΣ¬ΦΆΘ÷Ι, PNA”κRNA–Έ≥…ΒΡPNA2/RNAΡή“ΐΤπΖ≠“κΆΘ÷Ι[46]. ’β–©ΧΊΒψΕΦ «Ζ¥“εΙ―ΚΥή’ΥαΥυ≤ΜΨΏ±ΗΒΡ.
―–ΨΩ±μΟς, PNA ”κ mRNA ΫαΚœΡή”––ßΚΆΧΊΕ®ΒΊ“÷÷Τ Ζ≠“κ. Ι―ΚΥή’ΥαΒΡ÷ς“ΣΉς”ΟΖΫ Ϋ «Ά®ΙΐΒςΩΊ RNaseH ΒΡ ΫΒΫβΉς”Ο[47] . Εχ PNA ÷ς“Σ“άΩΩΈΜΉη”Αœλ, Αϋά®ΫαΚœ‘Ύ DNA “÷÷Τ RNA ΟΗΒΡΤτΕ·ΚΆ―”…λ[48] ΓΔΫαΚœ‘Ύ RNA ΉΣ¬Φ œύΙΊ«χ“÷÷Τ‘ΎΉΣ¬Φ«χΒΡΫαΚœΚΆΖ¥”Π[49] ΓΔΫαΚœ‘Ύ RNA ΒΡ Ζ«ΉΣ¬Φ 5'«χ“‘“÷÷Τ”κΚΥΧ«ΧεΫαΚœΜρΖ≠“κ[50] .
”…”Ύ»ΐΝ¥PNA2/DNAΚΆPNA2/RNAΫαΙΙΖ«≥ΘΈ»Ε®,“ρ¥ΥPNAΩ…“‘ΉςΈΣΖ÷Ή”Ά®ΒάΉη÷ΆΦΝ”Ο”Ύ“÷÷ΤDNAΜρRNAΒΡΟΗΖ¥”Π. ΕΥΝΘΟΗ «”…RNAΚΆΒΑΑΉ÷ Ήι≥…ΒΡΚΥΧ«ΚΥΥαΚΆΒΑΑΉΗ¥ΚœΈο, ΗΚ‘πΈ§≥÷»Ψ…ΪΧεΗ¥÷ΤΚσΕΥΝΘΡ©ΕΥ≥ΛΕ», ”ΟΉ‘…μ–·¥χRNA ΉςΡΘΑε, Ά®ΙΐΡφΉΣ¬ΦΚœ≥… DNA.PNA Ω…”κΧεΆβΒΡΕΥΝΘΟΗ÷– RNA ”––ßΚΆΧΊ“λ–‘ΒΡΫαΚœ[51], ’β“ΜΧΊ–‘‘ΎΆ®Ιΐ÷§÷ ΧεΫΪPNA ΒΦ»κΒΡ≈ύ―χΜυœΗΑϊ÷–“≤ΒΟΒΫΝΥ÷Λ Β[52]. ”…”Ύ‘ΎΕώ–‘÷ΉΝωœΗΑϊ÷–ΕΥΝΘΟΗ”–ΧΊ±πΒΡΜν–‘, PNAΩ…ΉςΈΣΕΥΝΘΟΗ“÷÷ΤΦΝ, ΩΣΖΔ≥…”––ßΒΡΩΙΑ©“©Έο.
PNA ”–ΩΙ HIV ΒΡΉς”Ο. ―–ΨΩœ‘ Ψ PNA Ω…“‘ΫαΚœ”Ύ ΡφΉΣ¬ΦΟΗΒΡ RNA ΡΘΑεΉηΑ≠ΡφΉΣ¬ΦΟΗΒΡΉς”Ο[53], “≤Ω…”κ RNA ΫαΚœ, Ά®Ιΐ“÷÷Τ HIV ≤ΓΕΨΗ¥÷ΤΒΡ±Ί“ΣΙΐ≥ΧΓΣΓΣΕΰ ΨέΧεΒΡ–Έ≥…, ά¥ΉηΑ≠ HIV ΒΡΗ¥÷Τ[54] .
Tyler Β»[55] Ήœ»±®ΒάΝΥΫωΆ®ΙΐΗΙ«ΜΉΔ…δ, Ζ«–ό ΈΒΡ PNA ΨΆΡήΙΜΉηΑ≠ΉΔ…δΫχ¥σΡ‘ΒΡ…ώΨ≠Ϋτ’≈ΥΊΒΡΉς”Ο, ±μ Ος PNA Ω…Ρή¥©ΙΐΝΥ―ΣΡ‘ΤΝ’œ. ΤδΚσ, Νμ“ΜΗω―–ΨΩ–ΓΉιΒΡ Bonnard Β»[56]Ά®Ιΐ σΗΙ«ΜΉΔ…δΩΙΑΔΤ§…ώΨ≠κΡ FF (NPFF) ΒΡ mRNA ΒΡΖ¥“ε PNA(»ΐ¥Έ, 10 mg/kg), Ιέ≤λΒΫΟςœ‘ΒΡ “÷÷ΤΩΙΑΔΤ§…ώΨ≠κΡ FF (NPFF)ΒΡΉς”Ο. œ‘ Ψ PNA ¥©ΆΗΝΥ ―ΣΡ‘ΤΝ’œ”κ NPFF ΒΡ mRNA ΫαΚœ. Adlerzc Β»[58]Ε‘≈ύ―χΒΡ σ–ΓΡ‘ΝΘœΗΑϊΚΆΤΛ÷ –«–ΈΫΚ÷ œΗΑϊΧεΆβ―–ΨΩΖΔœ÷, Έ¥–ό Έ PNA Ρή¥σ¥σœ¬ΒςΑΔΕζ¥ΡΚΘΡ§≤ΓΒΡ“ΜΗω÷¬≤Γ“ρΥΊΓΣΓΣ ΒμΖέΉ¥«ΑΧεΒΑΑΉ(amyloid precursor protein, APP). “‘…œΒΡ―–ΨΩ±μΟς, PNA Μα‘Ύ÷ΈΝΤ÷– ύ…ώΨ≠œΒΆ≥ΒΡΜυ“ρœύΙΊ≤Γ ÷ΔΚΆ―–ΨΩ÷–ΖΔΜ”÷Ί“ΣΉς”Ο.
»ΐΝ¥ΫαΙΙΒΡΗΏΈ»Ε®–‘“≤ Ι PNA Ω…”Ο”Ύ‘ΎœΗΑϊΡΎ–ό Η¥ΆΜ±δΒΡ DNA. Faruqi Β»[58]±®ΒάΝΥ PNA Ρή±ΜΈϋ ’»κ σ ΧεΡΎ, ΫαΚœ”ΎΜυ“ρΑ–Βψ, ΜαΒΦ÷¬ΒψΆΜ±δΓΔΒΞΦνΜυ»± ßΜρ ≤ε»κ, Τδ≈®Ε»÷Μ–η“Σ 10Θ≠7 mol/L. ¥ΥΆβ, ”…”ΎΫΪ PNA Ν§ ”Ύ÷ ΝΘΒΡΧΊΕ®≤ΩΈΜ, ≤ΜΜα”Αœλ÷ ΝΘΒΡ’ΐ≥Θ…ζάμΙΠΡή, “ρ ¥Υ PNA “≤Ω…“‘ΉςΈΣ“©Έο‘ΊΧε, ‘ΎΤδΙ«Φή…œΝ§Ϋ”κΡΓΔΒΑ ΑΉΓΔ“©Έο”Ο”ΎΜυ“ρ÷ΈΝΤ.
2.2 ’οΕœ
PNAΒΡΧΊ–‘ΡήΙΜΫβ ΆΥϋ‘ΎDNA’οΕœ…œΒΡ”Π”Ο. (1)PNA ΒΡΦνΜυ–ρΝ–Φχ±πΝΠΚΆΈ»Ε®–‘ΕΦΚΟ”ΎΙ―ΚΥή’Υα. (2)PNAΡήΙΜ“‘Ν¥«÷’ΦΒΡΖΫ Ϋ Ε±πΥΪΝ¥DNA.(3)PNA”κΜΞ≤ΙΚΥΥαΒΡΫαΚœΦΪ¥σΒΊΗΡ±δΝΥΒγ”Ψ«®“Τ¬ . “ρ¥ΥPNA ΡήΙΜΉςΈΣ’οΕœΧΫ’κ”Ο”ΎΦλ≤βΜυ“ρΆΜ±δΚΆ¥μ≈δΖ÷Έω.
άϊ”ΟΦνΜυΆξ»Ϊœύ≈δΒΡDNAΚΆ≤Μ»Ϊœύ≈δDNA”κΜΞ≤ΙPNA‘ΎΫαΚœΧθΦΰ…œΒΡ≤ΜΆ§, Ά®ΙΐΩΊ÷Τ‘”ΫΜΧθΦΰ, Βγ”ΨΡήΙΜΦλ≤βΒΫΝΫ’ΏΒΡ≤ν“λ[59]. ΗΟΖΫΖ® ”Ο”Ύ…Η―ΓΜυ“ρΆΜ±δ―υΤΖ.Igloi[60]ΫΪPNAΙΧΕ®”ΎΡΐΫΚ÷–, ‘ΎΒγ”Ψ÷–PNAΜαΕ‘ΜΞ≤ΙΙ―ΚΥή’Υα≤ζ…ζΉη÷ΆΉς”Ο, ≤Δ«“Ρή‘ΎΒγ”Ψ÷–Ϋχ––ΧΊ“λ–‘‘”ΫΜ. ’β÷÷ΖΫΖ®Ρή Β ±œ‘ ΨPNA”κΜΞ≤ΙDNA÷°ΦδΒΡ‘”ΫΜ«ιΩω.
Ι”Ο PCR Clamping ΖΫΖ®Ω…“‘Ζ÷ΈωΒψΆΜ±δ≤ΩΈΜ[61]. Α–ΚΥΥα PCR ά©‘ω «Φλ≤βΜυ“ρΆΜ±δΒΡ“ΜΗω÷Ί“Σ≤Ϋ÷η. PNA ”κ DNA ΒΡΫαΚœ≤ΜΡή±Μ DNA ΨέΚœΟΗ Ε±π, Ν¥≤ΜΡή―”…λ, ά©‘ω≤ΜΡήΫχ––. œύΖ¥ΤδΆΜ±δ–Ά‘ρΩ…“‘ΚήΚΟΒΊά©‘ω. Ι”Ο ’β“ΜΦΦ θΩ…“‘ΨΪ»ΖΒΡ÷Λ ΒΒψΆΜ±δΈΜ÷Ο.
“Μ÷÷≥ΤΈΣ”ΪΙβ‘≠ΈΜ‘”ΫΜ(FISH)≤βΕ®ΒΡΖΫΖ®ΚΆ PNA ΧΫ’κΦΦ θΫαΚœ, “―±Μ”Ο”ΎΑ©÷ΔΚΆΥΞάœ―–ΨΩ. PNA ΧΫ’κ”κ DNA ΧΫ’κΝΣΚœ, ”Ο”ΎΖ÷Έω X-ray ΒΦ÷¬ΒΡ»Ψ…ΪΧεΜΞΜΜ[62,63], “‘ΦΑ”Ο”Ύ»Ψ…ΪΧε“λ≥Θ’οΕœ[64] . PNA-FISH “≤±Μ”Ο”Ύ“Ϋ―ß ’οΕœΚΆΜΖΨ≥―υΤΖ÷–Φλ≤βΚΆΦχΕ®œΗΨζ[65,66], ’β÷÷Φλ≤βΖ«≥Θ ΩλΥΌΚΆΝιΟτ, ΒΪ≤ΜΡή«χΖ÷¥φΜνΒΡΚΆΥάΆωΒΡœΗΨζ.
2.3 Ζ÷Ή”…ζΈο―ßΙΛΨΏ
≥ΐΝΥ…œΟφΧαΒΫΒΡ“Μ–© Β―ιΦΦ θ,PNAΜΙΩ…”Ο‘ΎΖ÷Ή”…ζΈο―ßΒΡ–μΕύΖΫΟφ. PNA Ω…“‘Έ»Ε®ΚΆ–ρΝ–ΧΊ“λ–‘ΒΊ”κdsDNA –Έ≥…»ΐΝ¥ΒΡ«÷’ΦΫαΚœ, “ρ¥Υ PNA Ρή±ξΦ«÷ ΝΘDNA ‘ΊΧε[67]ΓΔΑ–±ξκΡ[68]Β». ’β÷÷±ξΦ«ΖΫΖ® «≤ΜΩ…ΡφΒΡ,ΒΪ»¥ «Ζ«Ι≤ΦέΦϋΫαΚœΚΆΓΑ…ζΈο―ß≥ΝΡ§Γ±ΒΡ±ξΦ«ΖΫΖ®.
‘ΎPCR ά©‘ωΙΐ≥Χ÷–, Φ”»κ“ΜΕΥΝ§”–”ΪΙβ–≈Κ≈ΜυΆ≈ΒΡΧΊΕ®–ρΝ–PNA, ”κDNAΑ–Ζ÷Ή”‘”ΫΜ, Ά®ΙΐΦλ≤βά©‘ωΙΐ≥Χ÷–ΥφΉ≈Έ¬Ε»±δΜ·, ‘”ΫΜΕχΗΡ±δΒΡ”ΪΙβ«ΩΕ», Ω…Ε‘ PCRΙΐ≥ΧΫχ––Φύ≤β[69,70].
3 ¥φ‘ΎΒΡ÷ς“ΣΈ Χβ”κΫχ’Ι
œ÷‘Ύ, PNA “―Ψ≠≥…ΈΣ“Μ÷÷Ζ«≥Θ”–”ΟΖ÷Ή”…ζΈο―ßΙΛ ΨΏ, ΒΪ“Σ≥…ΈΣ“Μ÷÷Μυ“ρ÷ΈΝΤ“©Έο, ”ωΒΫΚήΕύ’œΑ≠. άΐ»γ: Υ°»ή–‘≤νΓΔœΗΑϊΡΛΆ®ΆΗ–‘ΒΆΓΔPNA/RNA ≤ΜΡήΦΛΜν RNase HΓΔ”κΑ––ρΝ–ΫαΚœΈόΖΫœρ–‘Β». ”…”Ύ’β–©‘≠“ρ, –μΕύΙΪΥΨ ΚΆ―–ΨΩ–ΓΉι÷–ΕœΝΥΕ‘ PNA ΒΡ―–ΨΩ. ΒΪ «»‘”––μΕύ―–ΨΩ –ΓΉιΦα≥÷≤Μ–ΗΒΊΕ‘ PNA Ϋχ––―–ΨΩ, »ΓΒΟ“Μ–©Ϋχ’Ι, ≤ΩΖ÷ ΒΊΫβΨωΝΥ’β–©Έ Χβ.
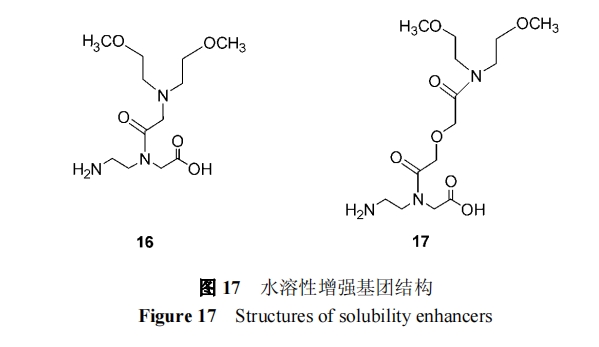
”…”Ύ PNA «Βγ÷––‘Μ·ΚœΈο«“”–Ή‘ΨέΦ·ΒΡ«ς Τ, “ρ ¥ΥΥϋΒΡΥ°»ή–‘Κή≤ν[71], ≤ΔΜαΥφΉ≈Ν¥ΒΡ‘ω≥ΛΚΆύ―Ώ ”κύΉ ύΛ±»ΒΡ‘ωΗΏΕχΫΒΒΆ[72] . ‘Ύ PNA ΒΡ C ΕΥ“ΐ»κάΒΑ±Υα[71]ΚΆ ‘ΎΙ«Φή…œ“ΐ»κ ÷–‘ΦΪ–‘ΜυΆ≈[20]ΕΦΡήΧαΗΏ PNA ΒΡΥ°»ή–‘. Gildea Β»[73]Κœ≥…Μ·ΚœΈο 16, 17(ΆΦ 17), Ν§”Ύ PNA ΒΡ N ΕΥ, “≤¥σ¥σ‘ωΦ”ΝΥ PNA ΒΡΥ°»ή–‘.
”…”Ύ ή PNA ΧεΡΎΉΣ‘ΥΡήΝΠΒΡœό÷Τ, ‘γΤΎΕ‘ PNA Ζ¥ “εΉς”ΟΒΡ―–ΨΩ, ÷ς“Σ «œό”ΎΧεΆβΈόœΗΑϊΧεœΒ. ΫΪ PNA ”κ ΒΑΑΉΒ»ώνΝΣ–Έ≥…«ΕΚœΈο, Ρή”––ßΒΡ Ι PNA ΆΗΙΐœΗΑϊΡΛ, ΉΣ‘Υ÷ΝœΗΑϊ ΓΔœΗΑϊΤςΜρœΗΑϊΚΥΡΎ. Muratovska Β»[74]ΫΪ PNA ”κ«Ή÷§–‘―τάκΉ”ΜυΆ≈ώνΝΣ, Ω…ΫΪ PNA ΉΣ‘ΥΫχ»κœΗ ΑϊΒΡœΏΝΘΧε÷–. Eriksson Β»[75]ΫΪ PNA ”κœΗΑϊΆΗΙΐ–‘ΕύκΡ ώνΝΣ, PNA Ω…“‘Υ≥άϊΆ®Ιΐ Escherichia coli ΨζΒΡΥΪ≤ψΡΛ, ΥΌ¬ ”κΩΙΨζκΡΒΡΥΌ¬ œύΒ±. Cutron Β»[76]ΫΪ PNA ”κΚΥΕ®ΈΜ–≈Κ≈κΡΚΆœΗΑϊΡΛΉΣ‘ΥκΡΝΣΫ”, Ω…“‘ PNA ΉΣ‘Υ÷ΝœΗΑϊΚΥ ΡΎ. ―τάκΉ”÷§÷ ΧεΑϋΙϋ «Νμ“Μ÷÷ΉΣ‘Υ PNA ΒΡ”––ßΖΫΖ®. ÷§÷ Χε≤Μ ΚœΉΣ‘ΥΒΞ“ΜΒΡ PNA, ΒΪΩ…“‘ΫΪ PNA ΚΆ DNA ΒΡ‘”ΫΜΈο[77]Μρ«ΕΚœΈο[78]ΉΣ‘Υ÷ΝœΗΑϊΡΎ.
Ι―ΚΥή’ΥαΒΡΖ¥“εΉς”Ο «Ά®ΙΐΦΛΜν RNase H ά¥ Βœ÷ ΒΡ, Εχ PNA ΒΡΖ¥“εΉς”Οά¥Ή‘”ΎΝΔΧεΉηΕœ. ΒΪ PNA ΚΆ DNA ΒΡ«ΕΚœΈο»¥ΡήΦΛΜν RNase H[79]. PNA ΒΡ¥φ‘ΎΩ…ΧαΗΏ …ζΈοΈ»Ε®–‘ΚΆ‘”ΫΜΒΡ«ΉΚΆΝΠ, Εχ DNA ‘ρΡήΧαΗΏΥ°»ή–‘ ΚΆΆΗΡΛΡήΝΠ.
PNA Ω…“‘ΤΫ––[3]ΜρΖ¥ΤΫ––[79] ΒΡΖΫ Ϋ”κ DNA ΚΆRNA ‘”ΫΜ, ’β”–Ω…ΡήΫΒΒΆΫαΚœΒΡΧΊ“λ–‘. Ήν≥θΒΡ―–ΨΩœΘΆϊΆ® Ιΐ“ΐ»κ ÷–‘ΜυΆ≈ΗΡ±δ PNA ΒΡ’β“ΜΧΊ–‘[21] . ΒΪΫαΙϊœ‘ Ψ, ÷–‘÷––ΡΒΡ“ΐ»κ÷ΜΡή¬‘”–ΗΡ…ΤΕχ≤ΜΆξ»ΪΫβΨω’β“ΜΈ Χβ. ΤδΚσ, ―–ΨΩ»Υ‘±Ε‘Ι«Φή¥χ”–ΜΖΉ¥ΫαΙΙΒΡ PNA Ϋχ––ΝΥ―– ΨΩ, ΫαΚœΒΡΖΫœρ–‘ Ε±π»‘÷Μ «”–ΥυΗΡ…Τ[31] .
Ψ≠Ιΐ °ΦΗΡξΒΡΖΔ’Ι, PNA ’ΐ÷πΫΞΉΏœρ≥… λ. 2002 Ρξ, ISIS ΙΪΥΨΙΚ¬ρΝΥ PNA ΒΡœύΙΊΉ®άϊ, ΉΦ±ΗΫΪΤδΖΔ’ΙΈΣΒΎ »ΐ¥ζΖ¥“εΙ―ΚΥή’Υα, ’β±ξ÷ΨΉ≈ PNA ≥…ΈΣ“©ΈοΒΡ«ΑΨΑ“― ÷πΫΞ±ΜΩ¥ΚΟ. ΗΡΫχ PNA ΒΡΧΊ–‘, ¬ζΉψ PNA ≥…ΈΣ“©ΈοΒΡ –η«σ, »‘ΫΪ «ΫώΚσ PNA ΒΡ―–ΨΩ÷ΊΒψ.
4 Ϋα¬έ
œ÷‘Ύ, κΡΚΥΥαΡή”κDNAΚΆRNAΧΊ“λ–‘ΫαΚœΦΑΜ·―ßΚΆ…ζΈο―ßΈ»Ε®–‘Β»ΧΊ–‘ ΙΤδ‘ΎΖ÷Ή”…ζΈο―ßΝλ”ρΒΟΒΫΝΥΙψΖΚΒΡ”Π”Ο, “≤¥χΕ·”κ÷°œύΙΊΒΡΜ·―ßΓΔΖ÷Ή”…ζΈο―ßΚΆ…ζΈοΦΦ θΝλ”ρΒΡΖΔ’Ι. ΒΪPNAΡήΙΜ≥…ΈΣΜυ“ρ÷ΈΝΤ“©Έο, »‘”–¥ΐ”Ύ‘ΎœΗΑϊΕ‘PNA ΒΡάϊ”Ο–ßΙϊΚΆΑ≤»Ϊ–‘ΤάΦέΖΫΟφ»ΓΒΟΗϋ¥σΒΡΫχ’Ι. ΈΣ¥Υ, ≤ΜΆ§ΫαΙΙΒΡPNAΒΞΧεœύΦΧ±ΜΚœ≥…≥ωά¥, ΩΦ≤λΤδΗς÷÷ΒΆΨέΧεΚΆ«ΕΚœΧεΒΡ…ζΈο―ßΧΊ–‘, “‘ΤΎΒΟΒΫΨΏ”–ΗϋΚΟΒΡ…ζΈοάϊ”ΟΕ»ΚΆ“©¥ζΕ·ΝΠ―ßΧΊ–‘ΒΡPNA,Έ“Ο«ΤΎ¥ΐΉ≈ΒΎ“Μ÷÷’φ’ΐΒΡPNAΜυ“ρ÷ΈΝΤ“©ΈοΒΡ≥ωœ÷.
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ